


Биосфера и ноосфера
Неизбежным следствием этого положения является ограничение всех величин, определяющих явления размножения организмов в биосфере. Должны существовать наибольшие числа неделимых, которые могут дать разные живые вещества. Эти числа – Nmax – должны быть конечны и характерны для каждого вида и расы. Скорости передачи жизни должны заключаться в точных и определенных пределах, которые не могут быть никогда превзойдены. Наконец, величины ∆ геометрических прогрессий размножения тоже имеют определенные пределы.
Эти пределы устанавливаются двумя проявлениями планеты: 1) ее размерами и 2) физическим заполнением пространства, в котором течет жизнь, жидкостями и газами – прежде всего свойствами газов и характером газового обмена.
§ 35. Остановимся на ограничении, вносимом размерами планеты. Влияние этих размеров мы видим на каждом шагу. Небольшие водоемы очень часто покрыты сплошь на своей поверхности плавающей на них зеленой растительностью. В наших широтах это очень часто зеленые ряски – разные виды Lemna. Поверхность воды представляет сплошной зеленый их покров без промежутков. Растеньица тесно сдвинуты, их зеленые пластинки заходят друг на друга; процесс размножения действует, но он замедлен внешним препятствием – прежде всего отсутствием места. Он проявляется только тогда, когда в зеленом покрове вследствие внешних разнообразных причин – гибели рясок или их уноса – образуются пустые промежутки водной поверхности. Они немедленно замещаются размножением. Очевидно, количество неделимых ряски, могущих поместиться на данной площади, определенно и находится в зависимости от размеров и условий их существования. Когда оно достигнуто, процесс размножения останавливается, задерживается внешним неодолимым препятствием. В каждом пруду создается своеобразное динамическое равновесие, аналогичное тому, какое в нем наблюдается при испарении воды с его поверхности. Упругость паров и упругость жизни механически аналогичны.
Другой всем известный пример представляет в картине природы жизнь зеленой водоросли – разных видов Protococcus, обладающей гораздо большей геохимической энергией, чем ряска. Она покрывает в благоприятных условиях сплошь без промежутков (§ 50) стволы деревьев. Дальше ей идти некуда; ее процесс размножения задержан; он возобновляется вновь, как только открывается возможность помещения новых неделимых Protococcus. Количество неделимых этой водоросли, могущих поместиться на площади дерева, строго определенно и не может быть превзойдено.
§ 36. Эти соображения могут быть целиком перенесены на всю живую природу и на область, доступную ее обитанию, – на поверхность нашей планеты. Наибольшее могущее существовать проявление силы размножения живого вещества определено размерами планеты и выражается в количестве неделимых, которые могут разместиться на площади, равной 5,10065 • 108 км2. Это количество есть функция густоты скопления организмов, возможной для их жизни.
Эта густота очень различна; для ряски или одноклеточной водоросли протококка она определяется только их размерами; другие организмы требуют гораздо большей площади (или объема) для жизни. Слон в Индии требует до 30 км2, овца на горных пастбищах Шотландии – около 105 м2, средний улей пчел – не менее 10–15 км2 (одна пчела – не менее 2 • 10-4 км2, т. е. 200 м2) среднего красного леса Украины, от 3000 до 15 000 неделимых планктона хорошо развиваются в 1000 см3 морской воды, 25–30 см2 достаточны для обычных злаков, несколько (иногда десятков) квадратных метров – для неделимых обычного нашего леса.
Очевидно, скорость передачи жизни зависит от возможной густоты хорошо живущей, не страдающей в своих проявлениях совокупности неделимых, от плотности живого вещества.
Не буду здесь останавливаться на очень еще мало изученной этой важной константе жизни в биосфере. Ясно, что наибольшая плотность сплошного покрова (типа ряски или протококка) или сплошного заполнения 1 см3 мельчайшей бактерией (§ 29) даст нам, если принять ее возможной для всех организмов, наибольшее допустимое для данного вида количество его неделимых в биосфере. Для получения этого числа необходимо принять плотность равной квадрату максимального изменения организма, т. е. его длины и ширины (коэффициент k1)[4].
§ 37. Ограничение размножения размерами планеты, неизбежная остановка процесса уже этим путем, помимо более глубокого влияния, оказываемого, как увидим, зеленой средой (§ 123), придает этому процессу очень своеобразные и важные черты.
Прежде всею, очевидно, есть предельное, одинаковое для всех организмов, наибольшее расстояние, по какому может распространяться передача жизни. Оно равно земному экватору, т. е. 40 075 721 м.
Во-вторых, для всякого вида или расы есть максимальное количество неделимых, которое не может быть никогда превзойдено. Это максимальное число получается при полном заполнении данным видом земной поверхности при максимальной возможной густоте его обитания. Это число, которое я буду дальше обозначать N и называть стационарным числом однородного живого вещества, имеет большое значение для оценки геохимического влияния жизни. Оно отвечает максимально возможному проявлению энергии данного однородного живого вещества в биосфере, максимальной его геохимической работе; скорость его достижения, разная для каждого организма, выражается скоростью V — передачи жизни.
Эта скорость – V — связана со стационарным числом следующей формулой:
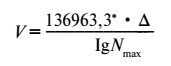
Очевидно, если скорость передачи жизни остается постоянной, ∆, характеризующая силу размножения (§ 32), должна уменьшаться, размножение организмов должно в данном объеме или на данной площади идти все медленнее и медленнее по мере того, как число созданных неделимых увеличивается, приближается к стационарному.
§ 38. Мы видим это явление в окружающей нас природе. Оно давно было замечено старыми натуралистами и ярко подчеркнуто около сорока лет назад точным наблюдателем живой природы К. Земпером (1888). Земпер отметил, что в небольших водоемах при всех равных условиях размножение организмов уменьшается по мере увеличения в них количества неделимых. Стационарное число не достигается или достижение его замедляется по мере приближения к нему количества создаваемых организмов; существует какая-то причина, может быть не всегда внешняя (§ 43), регулирующая процесс. Опыты Пирля и его сотрудников над мухой Drosophila и над курами (1911–1912) подтверждают это обобщение Земпера в другой среде.
§ 39. Скорость передачи жизни может давать нам ясное понятие о геохимической энергии жизни разных организмов. Она колеблется в больших пределах и находится в тесной зависимости от размера организма. Для самых мелких организмов, для бактерий, она, как мы видели, близка к скорости звука, т. е. к величине, равной 33 • 100 см/с. Для самых крупных, для крупных млекопитающих, она равна долям сантиметра – для индийского слона, например, V = 0,09 см/с.
Это крайние пределы. Между ними помешаются скорости передачи жизни для всех других организмов. Они находятся в явной зависимости от размеров организма и в более простых случаях (например, для организмов, форма которых приближается к шару) связь размеров организма с его скоростью v может быть уже сейчас математически выражена. Но существование определен ной математической зависимости всегда и везде в этой области несомненно и отвечает старинному прочному эмпирическому обобщению (§ 32).
§ 40. Скорость передачи жизни дает ясное понятие об энергии жизни в биосфере, о ее в ней работе, но оно недостаточно для ее определения. Для этого мы должны принять во внимание массу того организма, энергия растекания совокупностей которого в биосфере определяется скоростью V.
Выражение pV2/2, где р — средний вес организма[5], скорость растекания геохимической энергии которого равна V, даст нам выражение кинетической геохимической энергии живого вещества. Взятое по отношению к определенным площади или объему биосферы, оно может дать нам выражение той химической работы, которая в геохимических процессах этой площади или объема может быть произведена данным видом или расой организмов.
Уже давно мы имеем подходы к определению этим путем по отношению к определенной площади биосферы – гектару – части геохимической энергии живого вещества. Это делается при определении урожаев — количества с данной площади полезных человеку организмов или их продуктов.
В более полной форме оно выражается в количестве органического вещества, которое может быть создано размножением и ростом организмов на гектаре.
Хотя эти данные очень неполны и не охвачены теорией в достаточной степени, они привели уже к важным эмпирическим обобщениям.
Несомненно, количество создаваемого на гектаре органического вещества ограничено и теснейшим образом связано с той солнечной лучистой энергией, которую захватывает зеленое растение. Геохимическая энергия, собранная этим путем – размножением организмов, – есть измененная солнечная энергия.
Во-вторых, все более выясняется, что в случаях максимальных урожаев количества органического вещества с гектара почвы и гектара океана суть числа одного порядка, приближающиеся к одной и той же величине. Гектар суши охватывает ничтожный слой, не превышающий метров, гектар океана отвечает слою воды, захваченной жизнью, измеряемому километрами. Тождественность создаваемой в них энергии жизни, очевидно, указывает на освещение сверху как на ее источник.
Мы увидим, что, вероятно, это связано с характерным свойством почвы суши, скопляющей в себе концентрацию организмов, обладающих огромной геохимической энергией (§ 155). Благодаря этой концентрации энергии живого вещества тонкий слой почвы по своему геохимическому эффекту может сравниваться с огромной толщей моря, где центры жизни разжижены инертной массой воды.
§ 41. Кинетическая геохимическая энергия организма pV2/2, взятая по отношению к гектару, т. е. к 108 см2, выражается следующей формулой, где 108/k – количество организмов на гектар при достижении ими стационарного числа (§ 37), а k — коэффициент плотности жизни (§ 36):
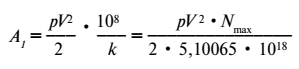
Чрезвычайно характерно, что эта величина для Protozoa есть величина постоянная. Для них всех выражение A1 принимает форму:
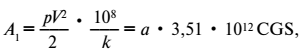
где а – коэффициент, близкий к единице[6].
Из этой формулы ясно, что кинетическая геохимическая энергия Protozoa определяется скоростью V, связанной с весом, размерами организма и темпом размножения Д. Отнесенное к ∆, V выражается следующей простой формулой:
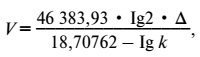
в которой числовые, постоянные для всех видов организмов коэффициенты связаны с размерами планеты (с площадью ее поверхности и с длиной ее экватора, причем величины выражены в сантиметрах и секундах)[7].
Из формулы скорости очевидно, что одни размеры планеты не могут объяснить действительно существующего предела для V и ∆.
Наибольшая нам известная величина V равна 33 100 см/с, а наибольшая величина ∆ – около 63–64.
Могут ли они идти дальше (что, как это видно из приведенных формул, возможно и при постоянстве кинетической энергии на гектар) или есть в биосфере условие, этому препятствующее?
Такое условие есть, и им является газовый обмен организмов, неизбежный и необходимый для их существования и, в частности, для их размножения.
§ 42. Организм не существует без газового обмена, без дыхания. Чем размножение идет быстрее, тем дыхание становится интенсивнее. По степени газового обмена мы всегда можем судить об интенсивности жизни.
В масштабе биосферы нам необходимо, конечно, иметь в виду не дыхание отдельного организма, а общий результат дыхания, необходимо учесть газовый обмен – дыхание всех живых организмов, обнять его как часть механизма биосферы.
В этом отношении давно имеются эмпирические обобщения, очень мало до сих пор обращавшие на себя внимание и не учтенные нашей научной мыслью.
Одно из них указывает, что газы биосферы те же, которые создаются при газовом обмене живых организмов. В биосфере существуют только они одни: О2, N2, СO2, Н2О, Н2, СН4, NH3… Это не может быть случайностью.
Затем весь свободный кислород биосферы создается на земной поверхности только благодаря газовому обмену зеленых организмов. Этот свободный кислород есть главный источник свободной химической энергии биосферы.
И, наконец, в-третьих, количество этого свободного кислорода в биосфере, равное 1,5 • 1021 г есть число того же порядка, как и количество существующего и с ним неразрывно связанного живого вещества, исчисляемого в 1020—1021 г. Оба эти исчисления получены вполне независимо друг от друга.
Эта тесная связь газов Земли с жизнью указывает с несомненностью, что газовый обмен организмов – и на первом месте их дыхание – должен иметь первостепенное значение в газовом режиме биосферы, т. е. являться планетным явлением.
§ 43. Этот газовый обмен – дыхание – определяет весь темп размножения; он ставит пределы для величин V и ∆.
Они не могут перейти через пределы, нарушающие свойства газов.
Я уже указывал (§ 29), что количество организмов, могущих существовать в одном кубическом сантиметре среды, должно быть меньше количества в нем газовых молекул, т. е. меньше 2,706 • 1019 (число Лошмидта). Если величина v будет больше 33 100 см/с, то для организмов, размеры которых меньше размеров бактерий (т. е. размеров меньшего порядка, чем п • 10-5), количество неделимых благодаря их размножению превысит 1019 в 1 см3. Очевидно, при неизбежном существовании обмена между газовыми молекулами и организмами количество поглощающих и выделяющих газовые молекулы организмов, соизмеримых с молекулами по своим размерам, должно было бы расти по мере уменьшения размеров организмов с все большей, в конце концов, невероятной быстротой.
По нашим современным представлениям, таким образом, приходим к физическому абсурду.
Если ограничение количества неделимых в кубическом сантиметре определяет наименьшие размеры организма и этим путем ставит максимальный предел для ∆ и V, то неизбежное постоянное соотношение между количеством неделимых и газовых молекул в данном объеме – явления дыхания — играет еще большую и постоянно проявляющуюся роль в явлениях размножения.
Дыхание, очевидно, регулирует весь этот процесс на земной поверхности, устанавливает взаимные соотношения между количеством организмов разной плодовитости, определяет, подобно температуре, величину ∆, которой может достигать данный организм в действительности; оно же определяет максимальную ∆, отвечающую размерам организма, не допускает достижения стационарных чисел.
В мире организмов в биосфере идет жесточайшая борьба за существование – не только за пищу, но и за нужный газ, и эта последняя борьба более основная, так как она нормирует размножение.
Дыханием определяется максимальная возможная геохимическая энергия жизни на гектар.
§ 44. Результат этого газового обмена и определяемого им размножения организмов огромен даже в масштабе биосферы.
Ничего аналогичного, даже в отдаленной степени, не представляет косная ее материя.
Ибо благодаря (размножению каждое живое вещество может создать новые любые количества живой материи. Вес биосферы нам неизвестен, но он составляет небольшую долю веса земной коры, до 16 км мощностью (§ 77), вес которой равен 2,0 • 1025 г. Равное весу коры количество вещества может быть силой размножения создано в ничтожное, не геологическое время, если только этому не препятствует внешняя среда.
Холерный вибрион и bacterium coli могут дать эту массу вещества в 1,60—1,75 суток. Зеленая диатомовая водоросль Nitzschia putrida – миксотрофный организм морской грязи, питающийся разлагающимися органическими веществами и в то же время захватывающий солнечный луч, – может дать 2,0 • 1025 г вещества в 24,5 суток. Это один из наиболее быстро размножающихся зеленых организмов, может быть, в связи с тем, что часть органических веществ он получает в готовом виде. Один из наиболее медленно размножающихся организмов – слон – может дать то же количество вещества в 1300 лет. Но что значат годы и столетия в геологическом, т. е. планетном, времени!
Мы должны к тому же иметь в виду, что при дальнейшем ходе времени новые массы, равные той же величине 2,0 • 1025 г, должны были бы получиться в несравненно более короткие сроки.
Эти числа дают нам понятие о тех силах, которые проявляются в явлениях размножения.
§ 45. Конечно, в действительности ни один организм не дает таких количеств.
Однако перемещения таких масс в биосфере силой размножения даже в течение одного года отнюдь не являются фантастическими, и даже в действительности они малы.
Эти числа не ирреальны. Мы действительно наблюдаем проявления жизни, им отвечающие, в окружающей нас природе.
Едва ли можно сомневаться, что жизнь в течение года путем размножения создает количества неделимых и отвечающие им массы вещества порядка 1025 г и, вероятно, в очень большое количество раз бóльшие.
Так, в каждый момент в биосфере существует п • 1020—п • 1021 г живого вещества. Это вещество вечно разрушается и создается главным образом не ростом, а размножением. Поколения создаются в промежутках от десятков минут до сотен лет. Ими обновляется вещество, охваченное жизнью. То, которое находится в каждую минуту в наличности, составляет ничтожную долю созданного в году, так как колоссальные количества создаются и разрушаются даже в течение суток.
Перед нами динамическое равновесие. Оно поддерживается трудно охватываемым мыслью количеством вещества. Очевидно, что даже в сутки создаются и разрушаются смертью, рождением, метаболизмом, ростом колоссальные массы живого вещества. Кто может измерить количества вечно создающихся и вечно гибнущих неделимых?
Это задача еще более трудного порядка, чем исчисление песчинок моря – задача Архимеда. Как исчислить живые песчинки, непрерывно меняющиеся в своем количестве с ходом времени?
Здесь одновременно скопляются и меняются бесчисленные неделимые в пространстве и во времени. Число их бывших и настоящих даже в течение одного человечески короткого промежутка времени превышает количество песчинок морского песка, несомненно, неизмеримо более чем в 1025 раз!
Зеленое живое вещество§ 46. По сравнению с силой размножения, с геохимической энергией живого вещества, массы его, находящиеся в каждый момент в биосфере, являются небольшими – 1020—1021 г.
Эти массы генетически связаны в своем существовании с зеленым живым веществом, единственным способным захватывать лучистую энергию Солнца.
К сожалению, наши современные знания не позволяют учесть, какую часть всего живого вещества составляет зеленый мир растений. Можно пока дать лишь очень приблизительное понятие о количественной стороне явления.
Нельзя утверждать, что количественно по своей массе зеленое живое вещество преобладает на всей поверхности Земли, но, по-видимому, оно преобладает на суше. В океане обычно считается, что количественно – по массе – преобладает животная жизнь. Если даже животная – гетеротрофная – жизнь преобладает в конце концов в массе всего живого вещества, то это преобладание не может быть очень велико.
Не разделяется ли живое вещество на две половины или почти на две половины по весу: на зеленое автотрофное и на его порождение – гетеротрофное? Ответить на этот вопрос мы не умеем.
Но, во всяком случае, несомненно, что уже одно зеленое живое вещество дает массы того же порядка – 1020—1021 г какие отвечают всему живому веществу.
§ 47. Строение этого зеленого трансформатора солнечной энергии на суше и в море резко различно. На суше преобладает травяная явнобрачная растительность, древесная составляет по весу значительную, может быть близкую ей, часть; зеленые водоросли и другие тайнобрачные, особенно протисты, отходят на задний план. В океане преобладают одноклеточные микроскопические зеленые организмы; травы, как Zostera, и большие водоросли составляют по весу небольшую часть растительной жизни; они сосредоточены у берегов и в более мелких местах, куда проникает солнечный луч; их плавающие скопления, как скопления саргассов в Атлантическом океане, теряются в общей безмерности морских пределов.
Зеленые метафиты преобладают на суше; из них наиболее быстро размножаются – обладают большей геохимической энергией – травы. Скорость передачи жизни древесной растительности, по-видимому, меньше. Зеленые протисты преобладают в океане.
Скорость V для метафитов едва ли превышает сантиметры в секунду, для зеленых протестов она достигает тысяч сантиметров, т. е. превышает в сотни раз силу размножения метафитов.
Это явление резко характеризует различие жизни моря и суши. Хотя в море зеленая жизнь, может быть, и менее господствует, чем та же жизнь суши, но общее количество зеленой жизни в океане благодаря его преобладанию над сушей на нашей планете по массе превышает растительность суши.
Зеленые протисты океана являются главными трансформаторами световой солнечной энергии в химическую энергию нашей планеты.
§ 48. Можно выразить разный энергетический характер зеленой растительности суши и моря в точных числах и иначе. Формула 2 n∆ = Nn (§ 32) дает нам приращение организма в сутки (α) при размножении; беря один исходный организм, мы имеем, для него (в первый день, когда п = 1):
2∆ – 1 = α
Откуда
2∆ = α + 1 и 2n∆ = (α + 1) n
Величина а есть постоянная для каждого вида; она определяет суточное приращение количества неделимых, сведенное к одному неделимому, т. е. указывает увеличение в каждые сутки одного неделимого.
Величина (α + 1)n, очевидно, определяет количество неделимых, создаваемое размножением в п-й день: (α + 1)n = Nn
Значение этих чисел видно на следующем примере. По М. Ломану, среднее размножение планктона (учитывая его гибель и поедание) может быть выражено константой α + 1, равной 1,2996. Та же постоянная для среднего урожая пшеницы во Франции равна 1,0130. Эти величины отвечают среднему идеальному суточному значению одного организма пшеницы и планктона после одних суток размножения. Отношение количеств неделимых планктона и пшеницы п первый день от начала размножения равно таким образом:
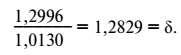
С каждым следующим днем это отношение будет расти согласно степени δ, т. е. будет в п-й день выражаться величиной δп.
Для 20-го дня величина δ равна 145,9, а для сотого дня количество неделимых планктона в 6,28 • 1010 раз должно быть больше количества неделимых пшеницы. В годовой оборот, после которого временно замирает развитие пшеницы, эта разница – δ365 – достигает астрономической цифры 3,1 • 1039. Конечно, при таком различии темпа размножения разница в весе взрослого травянистого растения суши, весящего сотни граммов, т. е. п • 102 г и микроскопического организма планктона, весящего немногие многомиллионные доли грамма (п • 10-6—п • 10-10 г), исчезает.
Зеленое организованное вещество моря достигает этого результата благодаря быстроте оборота своего вещества. Сила, в нем заложенная солнечным лучом, позволила бы ему создать в десятки дней, в 50–70 дней, а может быть и меньше, массу вещества, равную по весу земной коре (§ 44). То же предельное количество вещества могла бы дать травяная растительность суши в несколько лет – Solanum nigrum, например, в пять лет.
Необходимо иметь в виду, что эти числа не могут быть количественно сравнимы для выражения роли в биосфере зеленой травяной растительности и зеленого планктона.
Для такого сравнения надо их брать в одинаковые промежутки времени от начала процесса, причем различие быстро увеличивается с ходом времени. В то время как Solarium nigrum в 5 лет дал бы 2 • 1025 г вещества, зеленый планктон должен был бы дать в этот промежуток времени количества, которые трудно выразить понятными нам числами. В следующий, значительно меньший промежуток времени создания того же количества вещества травяной растительностью зеленый планктон дал бы еще большие, еще менее вообразимые числа.