
The Homing Instinct: Meaning and Mystery in Animal Migration
These behaviors get the animals to a good place (for overwintering or for reproduction). Like the long-range movements with specific endpoints on the map, homing to a good place is not always easily distinguished from moving out of a bad place. The behavior is a mechanism with deep evolutionary roots. Indeed, insect wings (and metamorphosis) may themselves have been an original adaptation for dispersal, to colonize temporary pools, animal carcasses, or other temporary resources. The first individuals to reach the resource won the competition to use it and multiply there, and these were more likely to be the ones that flew, and flew far and wide, rather than those that walked at random.
Wings and metamorphosis have lesser value in constant conditions. Some insects are able to respond in real time to the changes in conditions they experience (especially crowding), in that when they don’t “need” to disperse they either don’t grow wings (some aphids) or the muscles to power the wings are broken down and the amino acids from the protein are used instead to make more eggs (in some Hemiptera bugs). Often there are discrete “dispersers” versus “non-dispersers” in any given insect population, and the percentage of each depends on the quality of the home habitat and hence the relative cost/benefit ratio of moving versus staying.
Dispersing to “anywhere but here” generally applies to nonmigratory species that have no encoded or learned directions to go to but may have innate instructions to move in more-or-less straight lines rather than potentially going in circles in order to achieve distance. In Africa, dung-ball-rolling scarab beetles race away from their often thousands of competitors at a dung pile at night by using the swath of stars of the Milky Way galaxy as a reference. Swarms of insects feeding at dung and carcasses also attract predators, and as soon as they finish feeding, many distance themselves from those predators. I’ve observed blowfly larvae at animal carcasses keeping to almost perfectly straight lines in their getaway at dawn, by steering directly toward the direction of the rising sun. Mass movements sometimes observed in some rodents, such as lemmings and gray squirrels (as in 1935 in New England) following a population explosion after a superabundance of food, may be another example of dispersal to get to a better place, though not necessarily a predetermined one.
On the other hand, “true” migrants are able to utilize ideal conditions in two places, provided they vary predictably. Arctic terns, Sterna paradisaea, breed throughout the Arctic, then fly to Antarctica to escape winter when food availability declines and to arrive in spring and food again, a round-trip distance of nearly seventy-one thousand kilometers. Gray whales, Eschrichtius robustus, also feed in the Arctic in the summer but then travel eight thousand kilometers along the coastline to Mexico to bear their calves in warm waters.
Dispersers are not neatly differentiated from migrants, although the first commonly rely on passive mechanisms as opposed to the migrants, which move to specific goals by their own powers of locomotion. There are all gradations in between. Each case is unique, and there are thousands. Let’s look at more ways of getting to a good place, as represented by eels, a grasshopper, aphids, and ladybird beetles.
Eels. There are many species of eels, but the American, Anguilla rostrata, and the European, A. anguilla, the species with which we are most familiar, live most of their lives in freshwater ponds and lakes. For reasons that are not clear, though, they do not reproduce in their home areas. To the contrary, both disperse (or “migrate”) thousands of kilometers on a one-way trip to spawn and then die in the mid-Atlantic.
Just as birds and some insects use air currents, eels use water currents to help them leave their lifetime homes. After eels leave their freshwater homes and head for the ocean to spawn and die, their larvae then drift in the ocean currents for years. But eels’ dispersal behavior is anything but passive. Eels fatten up to prepare to leave their home haunts in the bottoms of freshwater lakes and streams. Before they head for the sea, they absorb their digestive tracts and transform themselves by greatly enlarging their eyes and turning silvery on their ventral side. The latter transformation produces counter-shading that reduces their visibility to predators below them in the open ocean waters.
The eels’ dramatic changes in behavior, morphology, and physiology that enable them to switch from living in freshwater habitat to open ocean highlight the operation of strong selective pressures. But why do they leave their homes in freshwater ponds where they grew up and lived most of their lives? Their one-way, once-in-a-lifetime migrations to their Sargasso Sea spawning grounds can’t be to find a better feeding ground, or to escape competition. However, I suspect what the behavior accomplishes superbly is that the adults, which are predators, do not come in contact with and feed on their own young. Is it a mechanism that has evolved because it reduced predation on themselves?
There are over six thousand publications on eels, but the life cycle of these economically important food fish is still murky and has a long history of speculation. For centuries, nobody ever saw a baby eel, and even now, their spawning has not been witnessed. Aristotle presumed that eels grew from earthworms. The first young of eels, transparent leaflike forms, were found in the open Atlantic Ocean. Gradually, as more of these leaflike creatures were collected, it was noted that they varied in size, and that the smallest ones were found south of Bermuda in the Sargasso Sea, which was therefore presumed to be the site of their origin, that is, the eel spawning area.
The eel larvae, after hatching in the waters of the Gulf Stream, drift north. Like plankton, they move at the whim of the prevailing oceanic current. As they grow from about five to six centimeters in a year, they take on a more eel-like form but remain transparent. They are by then able to swim and, presumably by scent, find and swim up a river. Unlike salmon, however, these transparent “glass eels,” as they are known at this larval stage, can have no specific home stream scent to follow, because they have experienced only the scents of the ocean.
The female glass eels at this stage, in early spring, migrate up rivers and streams along the East Coast of America. After two months in a river they grow to about ten centimeters. Now known as “elvers,” they are no longer transparent, and they enter lakes and become eels. These female eels live in lakes for eight years or more, fattening up (the males stay in the saline estuaries). When they achieve the right amount of fat, these females become sexually mature. Each develops a clutch of three to six million eggs, and then one fall the gravid females start their journey downstream back to the ocean, to the Sargasso Sea to spawn. Since the males don’t live in fresh water, somewhere in the ocean the females then apparently meet the males for fertilization.
As the Gulf Stream continues north beyond North America, the larvae of the European eel, which originate from the same apparent spawning area, the Sargasso Sea, as the American eels, continue their journey. Finally, in two or three years, they reach the coasts of Europe, where they then also seek rivers and streams. Migrating upstream, they become pigmented, and after growing to adulthood, they migrate back into the open Atlantic and make their sixty-five-hundred-kilometer return to the Sargasso Sea, where they spawn on average several million eggs, and then die. Only one of several million of them will make the return journey to grow to a reproducing adult in fresh water.
Grasshoppers. One of the best-known insect dispersers, the “migratory locust” (the grasshopper, Schistocerca gregaria), engages in some of the most spectacular mass movements in the animal kingdom. On the African continent, this species has been famous since biblical times. Swarms of the “locust” have blackened the skies, and as those in the vanguard settle onto the earth and consume every green thing where they land, the rest fly over them until they reach more green, while those behind then take flight and do the same, and so a horde of hundreds of millions moves along, stripping all vegetation in its path. Predators cannot put a dent in those hordes. Additionally, the migratory locust is distasteful to potential predators because when it migrates, it is not choosy about what it ingests and takes up toxins from poisonous plants, which it incorporates into its tissues. The grasshoppers’ bright red-orange and yellow coloration, like that of many insects including the monarch butterfly, reminds potential predators of its distastefulness.
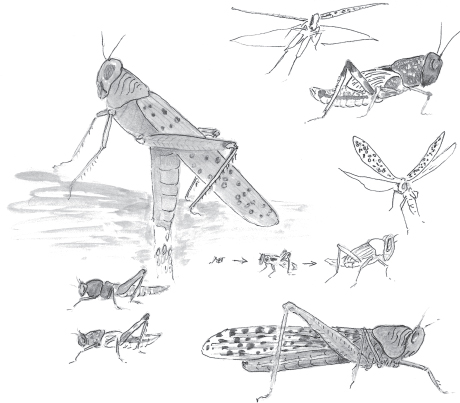
Nymphs and adults of the two phases of the migratory grasshopper
Although this distinctively colored grasshopper appears to arrive suddenly, it is often there all along, but in a different guise. It has a green solitary grass-fed form that blends in with its food and that is palatable to predators. For a long time scientists thought that the grasshopper “migrants” that appeared so suddenly were a unique species, one arriving from an unknown origin and heading for an unknown destination. Now we know that the migrants are a “phase” of a common species that changes its color, form, and behavior in response to crowding. Proof comes from experiments: to create these “migrants” from isolated individuals one takes a nymph (immature stage), puts it in a jar, and has a motor-driven brush tickle it continuously. The constant tickling mimics the crowding, which in the case of S. gregaria is the signal evolution has “chosen” to trigger the nervous system to alter the hormones that result in development into the restless migratory phase of a different color, wing length, and behavior. It is a good example that shows environment is “everything,” or from another perspective, it’s all about genetics.
Migratory-phase locusts are highly irritable and will jump up and follow a crowd flying over it. This behavior removes the grasshoppers from an area that is overpopulated and brings them to new land where conditions are conducive to feeding, egg-laying, and growth of their offspring. Although the grasshoppers could have no knowledge of where such a distant but good place might be, they migrate to it as if they do.
The grasshoppers reach a consensus. It is a sensible one, although it involves no thinking and no discussion. They simply fly up to join the crowd, which follows the prevailing winds. Eventually these winds meet air from an opposite direction and, when moist tropical air rises into cooler altitudes, rain precipitates out of the resulting clouds, depositing the grasshoppers to earth along with the rain. As the ground is watered and softened, the grasshoppers can shove their abdomens into the soil to lay their eggs. The new nymphs hatch just as new food starts to sprout. Their homing (or “dispersal”?), which has ended at this good place for them to reproduce, is now complete.
Aphids live in crowded “colonies” on plants into which they insert their mouthparts, much as mosquitoes puncture skin, except that they imbibe plant sap instead of blood and may stay plugged in at the same spot for most of their life spans. One might suppose they could not or would not migrate. But, like the migratory grasshoppers, they may travel possibly hundreds of kilometers. Nobody knows for sure how far; it depends on the prevailing winds.
Sedentary aphids already on good food do not leave to seek, or even require, mates. Instead, they switch to virgin births after a sexual migratory phase. Daughters then settle directly next to mother, and so on and on for many generations as the colony grows. And then, cued by the shortening of the days in the late summer and fall when the food supply runs out, the aphids’ offspring take a different developmental route. Because of changing day length and/or food, the nymphs on their final molt grow wings and become sexual. Frail and weak-winged they are, but an aphid is light and carried by the wind much like the seed of a dandelion or poplar tree, or a baby spider on a thread of silk. I usually see them in September when they appear like flecks of white lint floating erratically in the air.
To reach wind the aphids fly or are wafted up. Eventually, they don’t fight the wind but drift along and settle somewhere back down to earth. On their descent, assisted by their wings, they head toward anything colored light green. This color (unless they are tricked by pieces of green paper coated with sticky glue left by an insect physiologist studying them) is likely to be associated with their favorite food, fresh plant growth. After landing, perhaps because the chances of a mate arriving at precisely this one tiny spot of residence are remote, they switch back to virgin births and thus restart the cycle.
Ladybird beetles, the predators of aphids, similarly have adapted by migrating in a seasonal environment. In the western United States they migrate mainly on their own power from lowlands up into the Sierras, where they can in some locations be scooped up by the bucketful (generally to be sold to farmers and gardeners — to control aphids!). At the campus of the University of California at Berkeley, I often saw streams of them flying or being blown uphill in Strawberry Canyon by the campus when the grass was drying after the spring rains.
Ladybird beetles of some species migrate when reproduction must cease for the season. Despite the energy they expend for flight, they may migrate largely to save energy. It goes this way: As long as there are plenty of aphids to be had, both larvae and adult ladybirds don’t go hungry. Eventually, however, the green vegetation suitable for aphids disappears in the hot California summer, and so the aphids leave. Now the beetles’ resting metabolism kicks in as a significant liability. Resting metabolism for beetles is high at high body temperature but becomes almost negligible when they are torpid at the lowest body temperature tolerated, near or slightly below freezing. An elevated resting metabolism, month after month in the western states’ dry hot summer, would deplete both the beetles’ energy reserves and their body water. Without replenishment of food and water in that environment they would die. But by flying, with the aid of thermals, they are brought into upwelling air currents in the hills and then into the mountains where they reach cooler air. At this point, though, they do something different from the aphids: instead of being attracted to green, they are attracted to either red and/or the scent of each other. How else to explain that they crowd together into large groups in which they then overwinter? The advantage of their grouping behavior is not clear, but I suspect that it amplifies their noxiousness. (This is based on experience: ladybird beetles regularly come into my cabin to overwinter and quite often crawl into bed with me. I can vouch for the fact that they are noxious if not obnoxious, and several more so than one.)
The ladybird beetles arrive at a suitable place — a cool one — where they conserve their limited energy reserves during winter. The hypothesis that they home not just to an area, but also to a specific spot, is based on observations that my friend and colleague Dr. Timothy Otter has made in the Sawtooth Mountains near Stanley in Idaho. The beetles there were known by local ranchers to aggregate every fall in large numbers in specific rock cairns of decomposing granite in the hills above the valley floor. Otter, a biologist, wondered why the beetles aggregated there, in specific spots, and not in similar places nearby.
Since hibernation concerns adaptations related to temperature, Otter concentrated his efforts on unraveling the beetles’ temperature tolerance and compared it to the temperature at that site. But, curiously, there was nothing unique about the temperature of the specific site on “Ladybug Hill,” relative to other sites near it. Nor did the grouping of the beetles affect their temperature; the temperature of their aggregation was indistinguishable from ambient temperature. So, they didn’t aggregate to keep warmer than the environment there.
As already mentioned, the one thing all ladybug beetles do have in common is that they stink. The evolutionary significance of their aggregations is therefore likely the conventional one of other animals, namely, to advertise their noxiousness for protection from predators. Shrews and other predators may kill two or three victims and then spit them out, but they then learn to avoid them and so they do not continue eating the hundreds of thousands they would consume if they had found a bonanza of ladybirds. Given their safety of numbers, it is advantageous for any one beetle to join a group rather than to overwinter alone, because the chances that it will become a victim of a shrew’s educational process are reduced in more or less direct proportion to the group number. This rationale for large numbers of beetles, but of different individuals, massing repeatedly at the same location is not proven, but my colleague Daniel F. Vogt and I found that it applied to another noxious-smelling beetle, the whirligig (Gyrinidae) water beetles in Lake Itasca in Minnesota. These beetles there homed in on any existing groups of tens of thousands at dawn, after a night of foraging far and wide on the surface of the lake.
The second question is: How is an aggregation formed?
Insects have an impressive ability to home in on scent, and ladybird beetles could find the aggregation by following an odor plume. Memory cannot be involved in the ladybugs that Otter found year after year aggregating at the same site, which were generations removed from those of the previous year. The beetles arrive on site in September, stay there eight months, and in May return to the valley floor to feed, mate, and reproduce. Only their descendants, two or three generations later, could return to the tiny spot where they had hibernated.
The aphids’ rule of flying up toward the light, to then be dispersed by the wind, and then homing in on green when they settle could be a model of what happens in ladybird aggregation. Ladybirds are much stronger fliers than aphids, although they too are swept along in updrafts. But such drifting, while helping to account for their annual ascent from the lowlands into the mountains, does nothing to explain how tens of thousands of them end up under the same rock pile.
If the ladybirds home in on color, this could be tested, as the aphids’ homing in on food plants was tested — by leaving color targets at sites other than the traditional hibernaculum. But even if red color is an attractant (highly unlikely because the beetles aggregate under the rocks, not on them), that still would not explain their annual return to the same place in successive years. Do they smell their ancestors? Could thousands of smelly beetles piled up for eight months leave sufficient scent residue to serve as a marker that allows others to home in on the spot? If so, this idea could also be tested, by transferring an aggregation of beetles to overwinter at another physically similar place in the same general area as the old to see if a new traditional homing site for their mutual protection is created.
BY THE SUN, STARS, AND MAGNETIC COMPASS

Life has unfathomable secrets. Human knowledge will be erased from the world’s archives before we possess the last word that a gnat has to say to us.
— Jean-Henri Fabre
CHARLES DARWIN REFERRED TO THE ACCOUNT OF FERDINAND von Wrangel’s Arctic explorations, The Expedition to North Siberia, concerning how we home, quoting von Wrangel on how the Siberians oriented by “a sort of ‘dead reckoning’ which is chiefly affected by eyesight, but partly, perhaps, by the sense of muscular movement, in the same manner that a man with his eyes closed can proceed (and some men much better than others) for a short distance in a nearly straight line, or turn at right angles and back again.” Darwin compared a bird’s homing capability with that of people, but much less favorably, by telling how John James Audubon kept a wild pinioned goose in confinement, which at migration time became “extremely restless, like all other migratory birds under such circumstances; and at last it escaped. The poor creature immediately began its long journey on foot, but its sense of direction seemed to have been perverted, for instead of traveling due southward, it proceeded in exactly the wrong direction, due northward.” I’m not the least surprised at the behavior of the goose but all the more puzzled by our own orienting, which involves knowledge versus a feeling of “sense of direction.” I recall instances of waking up in “total” dark, “knowing” in my mind precisely how I am oriented relative to the room and hence the rest of my environment, but irked by “feeling” that I am in the precise opposite direction. It is then a struggle to get the two to agree, which happens only after some effort.
In Darwin’s time it was still supposed that humans had overall superiority over other animals. His then-hypothesis (later theory, and now fact) of evolution, which now binds us all as kin, was still revolutionary. Darwin found the goose’s behavior puzzling because he could not know that geese, cranes, and swans stay together in family and larger groups and that although the young by themselves do not know the correct migration route, they learn to know it from their parents which in turn learned it from theirs. Other birds have their migratory directions genetically coded, and they go strictly by “feeling,” since many of these have no knowledge because they migrate ahead of their parents.
We humans get lost easily. We would not get far without reference to landmarks, and I base that conjecture on (inadvertent) experiments. In one I was in long-familiar woods and got caught in a heavy snowstorm. Suddenly I got “turned around,” and it seemed as if all landmarks had in almost one instant been erased. But I kept going, trying to maintain a straight line by trusting my “sense of direction.” When I thought I had reached a place that I knew, where I should be going downhill, the landscape was instead sloping upward, and the brook I had expected to see was going in the “wrong” direction. At that point, knowing I was lost and no longer referencing to any signal, I backtracked in the snow and discovered that I had been walking in a circle, all the while thinking I was heading in the “right” direction. Yes, we can walk, a short way, in a relatively straight line with our eyes closed, by a process dubbed “path integration,” but my emphasis here is on “relatively.” Mice may do better. A friend told me of catching a Mexican jumping mouse in a live trap baited with peanuts. It had a kink in its tail, so he called it Crooked Tail, or CT for short. After he had released it several times, and it always returned for another snack of peanuts at the same source, he finally decided to “test its mettle” and released it exactly one kilometer away in thick brush and grass. The next morning CT was back for another snack. After release from two kilometers, though, it did not return. We don’t know, though, if this was due to failed navigation, finding a new food, a cost/benefit calculation, or a run-in with a coyote, owl, or weasel. On the other hand, when I failed to navigate, I was positive that I had been going in the precise opposite direction, which meant I had no sense of direction whatsoever, except that coming from visual landmarks from which I had constructed a map in my head.
When we do home, it is by maintaining a constantly updated calculation from at least two reference points, and the motivation to use them. We are innate homebodies, normally seldom displaced, so that in our evolutionary history there has been little need for a highly developed home-orienting mechanism. Simply paying attention to familiar landmarks suffices. Males on average may perform better than females in negotiating unknown territory, and it is posited that they, having been hunters traditionally, have a better “sense of direction.” But I doubt it. Learning, and especially attention, is hugely important for a presumed directional sense that can be developed to a high degree, as shown in some Polynesian seafarers living on isolated islands. But basically that involves being alert to more cues. These seafarers had been trained from near infancy to “read” the stars, the ocean waves, the winds, and other signs so that they may navigate over vast stretches of open ocean. But what a select few human navigators can accomplish with experience and with tools, many insects and birds do routinely as a matter of course, and with far greater precision over distances that span the globe.