
Палеонтология антрополога. Книга 2. Мезозой
Следующая великая группа рептилий – Lepidosauria; она тем более замечательна, что и поныне чувствует себя лучше прочих. Клювоголовые Rhynchocephalia (или клинозубые Spenodontida), ныне почти исчезнувшие, в триасе были многочисленнее. Planocephalosaurus и Brachyrhinodon taylori мало отличались от современной гаттерии, только вот их останки найдены в Техасе и Шотландии.
Как вы там, потомки?
Современная гаттерия Sphenodon punctatus (с природоохранными целями признано существование второго вида – Sphenodon guntheri) очень похожа на ящерицу, но это не ящерица. У гаттерий есть остатки хорды, брюшные рёбра, венозный синус в сердце, нёбные зубы и функционирующий третий глаз с хрусталиком, но нет копулятивного органа, барабанной перепонки и стремечка, а полость среднего уха у неё заполнена жиром. Живут гаттерии только на маленьких островках вокруг Северного острова Новой Зеландии.
Ещё лучше дело сложилось у чешуйчатых Squamata. Megachirella wachtleri из среднего триаса Италии выглядела довольно банально – именно так, как полагается великому предку всех ящериц. Гораздо оригинальнее планирующие Kuehneosauridae – американский Icarosaurus siefkeri и европейский Kuehneosaurus latus, чьи невероятно удлинённые ребра образовывали «крылья» по бокам тела, как ранее у Weigeltisauridae, одновременно у Mecistotrachelos, позже у Xianglong и в наше время у Draco. Самое интересное, что все эти группы не происходили друг от друга, а развили полёт на рёбрах совершенно независимо.

Упомянутый Mecistotrachelos apeoros из карнийского века Виргинии внешне был очень похож на кюнеозаврид – тоже «ящерка», планирующая на спицеобразных рёбрах, но представляет совсем другую – и гораздо более знаменитую группу – Archosauromorpha.
Архозавроморфы оказались особенно разнообразными диапсидами. В триасе возникла целая куча вариантов архозавроморфов: Protorosauria, Trilophosauria, Fenestrasauria, Rhynchosauria и Archosauria. Впрочем, такое деление далеко не единственное возможное, палеонтологи предложили массу альтернативных схем.
Кроме помянутых групп архозавроморфов, известно много отдельных родов и видов, которые не находят себе прочного места в системе. Например, полуводные саблезубые двухметровые Vancleavea campi и Litorosuchus somnii были похожи на талаттозавров, но, видимо, не были им родственны. Тело этих животных было запаковано в прочный чешуйчатый панцирь, причём в разных местах тела чешуи отличались размером и формой; особенно оригинальны высокие плоские пластинки, образовывавшие гребень по верхнему краю хвоста у ванклеавеи. Впрочем, бронированность, вероятно, не очень мешала подвижности.
Одна из примитивнейших групп архозавроморфов – Protorosauria (они же Prolacertilia, или Prolacertiformes), а её примитивнейшие представители – южноафриканская раннетриасовая Prolacerta broomi и индийская среднетриасовая Pamelaria dolichotrachela – выглядят уже не стандартно, как длинноногие и длинношеие ящерицы с маленькими головками. Похожи и Macrocnemus, и крошечный – всего 14 см – Cosesaurus aviceps.
Работа над ошибками
Prolacerta внешне настолько похожа на ящериц, что некоторое время считалась их прямым предком, за что и получила своё название. Стройные макрокнемус и казезавр, между прочим, выглядят так, как в теории должен был бы выглядеть предок птерозавров, хотя у того, надо думать, передние лапки должны бы быть длиннее задних, тогда как у проторозавров соотношение обратное.
Шея продолжала расти и у китайского Dinocephalosaurus orientalis вытянулась до 1,7 м из 3,5 м общей длины! Тело диноцефалозавра было тонкое и длинное, ножки коротенькие. Этот ящер замечателен не только пропорциями, но и формой головы: плоской, широкой, длинной, прямоугольной, со вздёрнутым носом и огромными острыми клыками. По всей видимости, это был рыбоядный обитатель вод. Совершенно сенсационна находка фрагментов скелета диноцефалозавра с косточками детёныша внутри. Речь не идёт о каннибализме, эти чудесные рептилии были живородящими! Было ли это связано с жизнью в воде или другими сложностями, понять трудно.
Маленькая тонкость
Живорождение диноцефалозавра как минимум значит хромосомное определение пола. У современных крокодилов пол детёныша определяется температурой, при которой находится яйцо. Например, у миссисипских аллигаторов Alligator mississippiensis при 30 °C вылупляются самки, а при 34 °C и выше – самцы. Сама же температура в немалой степени зависит от расположения гнёзда. Подобным образом дело обстоит и у многих черепах, ящериц и гаттерий. Но при живорождении температура матери вряд ли будет столь существенно различаться, причём долгое время, так что пол, скорее, будет зависеть от специальных хромосом, хотя ясно, что генов триасовых животных никто никогда ещё не видел.
Как бы ни был вытянут диноцефалозавр, в номинации «живая удочка» однозначно побеждает Tanystropheus, у которого из максимальной длины 5–6 м шея занимала 3–3,5 м. Шейные позвонки были чрезвычайно удлинёнными, из-за чего, правда, шея не была очень уж гибкой, особенно плохо она гнулась по вертикали. Но удилищу и не надо быть слишком уж эластичным: танистрофеусы сидели на берегах, не спеша макая свои длинные шеи в воду и вылавливая оттуда рыбу, чешуя которой найдена в районе желудка некоторых скелетов. Чтобы невероятная шея не перевешивала, танистрофеусы имели толстый зад и длинный хвост, который, кстати, не был уплощён и вряд ли мог работать веслом. Вероятнее всего, огромные шеи диноцефалозавров и танистрофеусов возникли независимо, хотя очевидно, что предрасположенность к этому была уже у первых проторозавров. Примечательно, что зубы детёнышей отличаются от зубов взрослых своей трёхвершинностью, вероятно, маленькие танистрофейчики были насекомоядными и лишь с возрастом переходили на рыбную диету. Для рептилий такая смена вкусов – обычное дело, но смена зубов нестандартна.
Работа над ошибками
В 1852 г. кости Tanystropheus conspicuus были описаны как хвостовые позвонки динозавра, в 1886 г. новые находки были определены как фаланги птерозавров Tribelesodon longobardicus. Лишь в 1929 г. обнаружение полного скелета расставило всё на свои места: вытянутые косточки оказались не хвостовыми позвонками и уж тем более не пальцами, а шеей архозавра-рекордсмена.
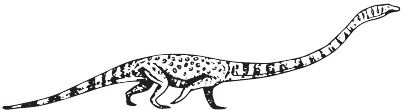
Tanystropheus
Langobardisaurus pandolfii тоже имел удлинённые шейные позвонки, но самих позвонков было немного. Его голова была непропорционально большой, ноги – тонкими и длинными, кисти и стопы – маленькими, а на хвосте первые хвостовые позвонки имели очень длинные поперечные отростки, а дальше тянулась хлыстоподобная нить вдвое длиннее тела. В длину эти странные животные достигали от силы полуметра. В передней части челюстей зубов не было, далее следовали трёхбугорчатые зубы, а завершался ряд огромным длинным плоским моляроподобным зубом. Ещё один родственник танистрофеуса – Tanytrachelos ahynis – имел огромные задние лапы и маленькие передние, его шея была длинной, но до своего знаменитого полутёзки ему было всё же далеко. Особая фишка танитрахелоса – специфические постклоакальные кости за тазом под хвостом. То ли они поддерживали сумку для вынашивания яиц, подобно сумчатым костям сумчатых млекопитающих, то ли служили для прикрепления особых мышц – непонятно.
Некоторые проторозавры пошли совсем другими путями. Ящерицеподобные Drepanosauromorpha – сплошные уникумы, просто победители конкурса «мы ищем таланты». Они освоили деревья и развили для этого массу странных особенностей. Самый древний представитель – Kyrgyzsaurus bukhanchenkoi – найден в Киргизии, а прочие представители – в Европе и Северной Америке. Крошечный – всего 15 см – итальянский Vallesaurus имел короткую мелкозубую мордочку, кривенькие цепкие лапки, но ещё простого строения, увеличенные остистые отростки передних грудных позвонков, а также хвостик с удлинёнными верхними и нижними отростками на позвонках в передней половине и тонким, заворачивавшимся спиралью концом. Вероятно, таким хвостом, похожим и на весло, и на крючок, можно было цепляться за ветки, как это делают цепкохвостые обезьяны и кинкажу.
Развитие дрепанозавроморфов можно видеть на итальянском Megalancosaurus preonensis, выраставшем до 25 см: его вытянутая треугольная голова с крошечными треугольными зубками была удивительно похожа на птичью, глаза были направлены вперёд, передние лапки были увенчаны кривыми цепкими пальцами с загнутыми когтями, причём два были противопоставлены прочим трём, на задних большой палец, снабжённый копытцем, был противопоставлен четырём прочим когтистым, остистые отростки передних грудных позвонков были увеличены и странно загнуты, а на третьем и четвёртом грудных позвонках вообще срослись, последний же позвонок загнутого хвоста превратился в крючок для цепляния за ветви. Очень длинные лапы позволили предположить, что между ними была натянута летательная перепонка, как у летяги, хотя прямых доказательств этому нет.
Без сомнения, всех превзошёл Drepanosaurus unguicaudatus, несуразный во всём. Его передние лапы не похожи ни на чьи: плоская гребнистая локтевая кость торчала под прямым углом к укороченной лучевой, их концы не соединялись между собой, а огромные – больше лучевой – кости запястья образовали какую-то непотребную конструкцию, если на что и похожую, то на «трёхзвенное» или даже вообще дополнительное предплечье. Из пяти пальцев первый и три последних были маленькими и тоненькими, а второй развился в огромный штырь с гигантским кривым когтем. Самые смелые палеонтологи даже предположили, что такая удивительная рука-крюка предназначалась для расковыривания насекомьих гнёзд типа термитников; правда, существование термитов в триасе пока не доказано, но теоретически что-то подобное уже могло иметь место. Задние лапы дрепанозавра тоже были вооружены огромными кривыми когтями. На загривке дрепанозавра красовался здоровенный гребень из сросшихся отростков позвонков, надо думать – для крепления мощнейших плечевых мышц. Хвост был расширен за счёт удлинённых верхних и нижних отростков позвонков, загнут дугой, а самое главное – нёс костяной крючок на кончике, как у скорпиона. Видимо, подвешивание на ветвях дрепанозавр развил в полном совершенстве.

Drepanosaurus unguicaudatus
Двенадцатисантиметровый Hypuronector limnaios из Нью-Джерси выглядел чуток попроще – без экстремальных пальцев и с совсем небольшим горбиком на загривке. Зато хвост гипуронектора был поистине красив: в виде огромного – целых 6 см! – пера с длиннющими нижними отростками на хвостовых позвонках. Зачем гипуронектору было такое украшение – вопрос. То ли хвост позволял прикидываться листом, то ли, как предполагают некоторые, был рулевой частью летательного оснащения, тогда как между длинными ножками могла бы быть натянута летательная перепонка. Впрочем, как и в случае с мегаланкозавром, это только гипотеза.
Стремление к полёту обуревало и других архозавроморфов. Чудеса продолжались. Sharovipteryx mirabilis из Киргизии и Ozimek volans из Польши были бы очередными тощими длинноногими «ящерками», если бы не летательная перепонка, растянутая на задних ногах. Крылья и всяческие иные летательные плоскости возникали в процессе эволюции не раз, но только шаровиптериксы умудрились сделать это на основе задних ног! Летательная перепонка занимала пространство между палочковидными ногами и основанием хвоста и не соединялась с крошечными передними лапками. Судя по мелким зубкам, озимек был насекомоядным, так что планирование было полезно как для спасения, так и для нападения. Конечно, кроме «ножных крыльев», были и другие странности. Так, у озимека шейные позвонки были крайне удлинены, напоминая вариант танистрофеусов, коракоид срастался с грудиной в единую грудную пластину, лопатка выглядела как широкая полулунная пластина, а на стопе среди тонких вытянутых прямых косточек резко выделялась толщиной, краткостью и изгибом пятая плюсневая кость. Вероятно, она служила опорой при базировании на ветках.
Маленькая тонкость
Каких только летательных приспособлений не существует! Над водами морей парят на растопыренных плавниках некоторые кальмары – крылорукий Sthenoteuthis, птицекрылый Ornithoteuthis и короткопёрый Illex, а также целое семейство летучих рыб Exocoetidae. Особенно богата на умельцев-планеристов Юго-Восточная Азия. Веслоногие лягушки Rhacophorus планируют на перепонках, натянутых между пальцами рук и ног. Лопастехвостые гекконы Ptychozoon имеют не только перепонки на пальцах, но и расширенное тело и плоский хвост. Украшенные древесные змеи Chrysopelea из Юго-Восточной Азии летают вовсе без ног, распластывая всё своё длинное тело. Среди млекопитающих тем же заняты шерстокрылы Cynocephalus volans и Galeopterus variegatus, сумчатые летяги Petaurus, гигантские летяги-тагуаны Petaurista, шипохвостые летяги Anomalurus и наша родная летяга Pteromys volans.
Активный полёт – куда более сложное занятие. Из беспозвоночных его освоили только крылатые насекомые, а из позвоночных – лишь птерозавры, конфуциусорнисы, энанциорнисы, веерохвостые птицы и летучие мыши. Что характерно, по крайней мере птерозавры появились всё в том же триасе. Выдвинуто даже предположение, что шаровиптериксы явились предками птерозавров, однако это крайне сомнительно: шаровиптериксы парили пассивно на ножных перепонках, а птерозавры – активно порхали на ручных.
Ещё одно диво из Киргизии – Longisquama insignis – загадочная диапсида, лишь условно относимая к архозавроморфам, истинная дракошка. На её спине красовался ряд длиннющих – до 29 см, вдвое длиннее всего тела – чешуй. С момента открытия первого отпечатка лонгисквамы в 1969 г. учёные спорят о том, как были устроены и зачем вообще возникли эти «перья». Не вполне ясно, располагались ли они в один или в два ряда, имели ли они настоящие бородки, как у современных птиц, или были сплошными (у примитивных меловых птиц они, кстати, тоже бывали сплошными). По средней линии «перьев» шёл полый стержень, а «опахало» было асимметричным, как у летающих птиц. По одной из реконструкций, «перья» могли раскладываться вбок подобно крыльям и служили для полёта, по другой – не имели никакого отношения к планированию, складывались назад и служили либо для мимикрии, либо для брачных демонстраций. Вторая версия кажется более убедительной, по крайней мере, на самом лучшем отпечатке никаких следов второго ряда не видать, а «бородки» всё же не были разделены, так что опахало было сплошной мембраной. Мы не знаем этого наверняка, но, если удивительные чешуи были порождением полового отбора, можем предположить, что они были ещё и ярко окрашены.

Longisquama insignis
Кроме «перьев», лонгисквама удивительна и остальными пропорциями: её треугольная глазастая головка по длине почти равна пятисантиметровому тельцу (впрочем, задняя половина скелета не сохранилась), а передние лапки – ещё длиннее, причём размеры плеча, предплечья и кисти почти одинаковы, лопатка длинная и тонкая, почти как у птиц, ключицы, вероятно, срастались в единую вилочку. Шейных и брюшных рёбер нет – вроде бы для пущей воздушности, однако кости не имели воздухоносных полостей. Всё тело было покрыто длинными армированными чешуйками.
Лонгисквама, шаровиптерикс, озимек и некоторые другие нестандартные триасовые архозавроморфы иногда объединяются в группу Fenestrasauria, хотя её статус очень шаток.
Одни из самых обычных животных триаса – Rhynchosauria. На удивление, они крайне непопулярны среди художников, существуют считанные реконструкции их облика, а на масштабных панно триасовых пейзажей Вы никогда не увидите стад ринхозавров, хотя в реальности они были аналогом современных некрупных копытных, типа свиней или овец, основным мясом для хищников, наряду с дицинодонтам. Ринхозавры были низкими коротконогими зверушками обычно меньше полутора метров длиной (впрочем, некоторые вырастали и до пяти метров), но с крупной и очень странной головой. Примитивные раннетриасовые Noteosuchus colletti и среднетриасовые Mesosuchus browni ещё имели маленький клюв с зубами, но более продвинутые Rhynchosaurus articeps и Hyperodapedon обзавелись уже здоровенным узким беззубым клювом в передней части морды. Череп непрерывно расширялся в задней части, пока у гиперодапедона ширина не превзошла длину. Задние и нёбные зубы слились в длинные и широкие зубные пластины-щётки с массой пупырышек, которыми ринхозавры перетирали жёсткие растения. Учитывая низкорослость и форму клюва с зубами, предполагается, что ринхозавры ели корни, выдёргивая их из земли. Как ни странно, активное жевание сочеталось с отсутствием вторичного нёба. Вместе с тем, наружное носовое отверстие из-за сужения передней части морды стало непарным, прямо как у млекопитающих, что крайне нетипично для рептилий.
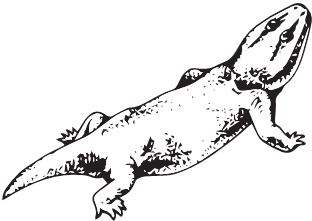
Rhynchosaurus articeps
Примечательна география ринхозавров: гиперодапедон известен в Южной Америке, Африке, на Мадагаскаре, в Индии, Северной Америке и Европе, то есть, проще говоря, везде. Неспроста в разное время разные исследователи описали массу родов и видов – Cephalonia, Isalorhynchus, Macrocephalosaurus, Paradapedon, Scaphonyx, Stenometopon и Supradapedon, которые при ближайшем рассмотрении оказываются одним и тем же гиперодапедоном.
Маленькая тонкость
Остатки ринхозавров найдены во многих местах, в том числе в Шервудской формации, названной в честь того самого Шервудского леса в Ноттингемшире, где разбойничал Робин Гуд. Знаменитый грабитель богачей не знал, что бегает по триасовым отложениям с ценнейшей палеонтологией.
В конце триаса из каких-то проторозавров появились Trilophosauria – в целом примитивные архозавроморфы, но с уникальной зубной системой. Это были приземистые тяжеловесные длиннохвостые создания 2,5 м длиной, ползавшие на раскоряченных длиннопалых когтистых лапах. Необычным было то, что у трилофозавров закрылось нижнее височное окно. У примитивного Variodens inopinatus передние зубы были коническими, а задние – резко расширенными гребнистыми трёхбугорчатыми «молярами». Дальше пошёл Trilophosaurus: его небольшая укороченная голова с высоким затылком – для увеличения жевательной мускулатуры – была снабжена спереди роговым клювом, а задние зубы стали ещё более узкими трёхвершинными гребнями, причём верхние и нижние плотно смыкались. Видимо, такая зубная система была предназначена для пережёвывания растений, причём жевательные движения происходили скорее спереди-назад, а не поперечно. Совсем странным был Teraterpeton hrynewichorum, чья морда вытянулась в длинный узкий беззубый клюв (в задней части челюстей и нёба мелкие зубы присутствовали), а ноздри растянулись на половину длины черепа.

Что сказать, триас – воистину время чудес. Но чудеса по определению не могут быть повторяющимися, слишком частыми и долгими. Да к тому же в любом сказочном лесу должны быть свои чудовища.
Особенно успешными архозавроморфами оказались собственно архозавры Archosauria. Именно они в итоге вытеснили всех и вся, в буквальном смысле съели всех удивительных родственников, да и вообще современников. В триасе архозавры пёрли напролом по принципу «всех убью – один останусь». И ведь им это почти удалось! Весь оставшийся мезозой прошёл под тяжкой пятой архозавров.
Залогом бешеного успеха архозавров стали всего две особенности: мощные зубастые челюсти с текодонтными зубами и вертикальное расположение задних ног под телом. Первое позволило кусать всех без всяких лишних заморочек с гетеродонтностью и жеванием, а второе – догнать кого угодно, чтобы его, собственно, покусать.
Маленькая тонкость
Зубы рептилий крепятся к челюстям тремя основными способами. Акродонтные зубы нарастают поверхностно или крепятся к наружной стороне челюсти, а потому непрочно; такой вариант типичен для рыб, амфибий и гаттерий. Плевродонтные зубы вырастают с внутренней поверхности челюсти, прилежа к ней на некотором протяжении, но не погружаясь в кость; они обычны у ящериц. Текодонтные погружены в специальные ячейки – альвеолы, а их основание – корень – закреплено соединительной тканью. Архозавры обрели текодонтные зубы, что дало им возможность кусать смелее и сильнее – зубы перестали вылетать по любому поводу.
Конечности у подавляющего большинства рептилий широко расставлены в стороны, так что тело большую часть времени лежит на пузике. Даже при беге пресмыкающимся часто приходится волочить брюшко и хвост по почве, что, во‐первых, дало им название, а во‐вторых, оставляет характерные изгибающиеся следы. Архозавры были едва ли не вторыми после зверообразных, кто надёжно поднялся над землёй. Только, в отличие от тероморфов, архозавры сделали ставку именно на задние ноги, а потому быстро и неоднократно стали подниматься на две ноги. Предпосылки к этому видны даже у проторозавров, для некоторых из которых, кстати, предполагается периодическая двуногость. Но архозавры решили проблему хорошо и надёжно. Бег архозавров ускорился, и никто уже не мог от них скрыться!
Современные архозавры – крокодилы – имеют едва ли не самый примитивный для архозавров вариант расположения ног. Возможно, именно архаичность позволила им пережить всех своих продвинутых родственников. Но в минуты особой бодрости (когда, например, приспичило догнать кого-то особо шустрого) даже крокодилы способны проявить свою архозавровую сущность – поднимаются на выпрямленных ногах и скачут галопом, да к тому же способны подниматься на две ноги. Правда, они делают это редко и неохотно, но наследие триасовых предков всё ещё живо в их ногах.
Особая прыткость в тёплом триасе не требовала особой теплокровности. Смотря на крокодилов, можно предположить, что уже в триасе развилось четырёхкамерное сердце и диафрагма, но на их же примере видно, что эти усовершенствования – не залог и не обязательное следствие теплокровности. Впрочем, обмен всё же не мог не ускориться. А побочным эффектом усиленного обмена и необходимостью при более шатком и сложном способе передвижения стало развитие мозга и усложнённое поведение.
Архозавры не слишком разнообразны, но это не облегчает их классификации. В классической схеме выделяются текодонты Thecodontia, крокодилы Crocodylia, птерозавры Pterosauria, ящеротазовые динозавры Saurischia и птицетазовые динозавры Orhithischia, но существует масса альтернативных схем, в которых текодонты расформировываются на кучу слабосвязанных группок, и даже динозавры то подразделяются на несколько отрядов, то перегруппировываются в совершенно невообразимом порядке. Апофеозом новаторства является включение в архозавров птиц Aves. Эти несомненные потомки архозавров по кладистической логике обязаны быть в них включены, хотя, с точки зрения орнитологов, такое объединение имеет очень мало смысла.
Маленькая тонкость
Существуют несколько способов представления родства живых существ. Самое модное и практически полностью победившее нынче направление – кладистика. Ключевая идея кладистики – классификация должна максимально отражать филогению, то есть порядок возникновения одних групп от других. Есть несколько вариантов кладистики, в самом жёстком и каноничном масштаб возникающих у потомков отличий от предков вообще не играет никакой роли, во внимание принимается лишь последовательность ветвления эволюционных линий. Соответственно, и названия можно давать лишь всем потомкам какого-то предка (монофилетической группе), но нельзя особо выделять лишь одну часть потомков, игнорируя другую. Исходя из этой логики, многие традиционные группы теряют свой смысл. Например, если птицы ответвились от некой части архозавров, а млекопитающие – от части зверообразных рептилий, то, при признании птиц и млекопитающих самостоятельными группами, нет никаких «рептилий» (поэтому многие современные зоологи очень болезненно реагируют на словосочетание «зверообразные рептилии») и даже нет динозавров (отчего другие столь же рьяно не забывают повторять «птицы – это тоже динозавры!»). При последовательном применении этой методики на планете нет даже прокариот (если признать самостоятельность эукариот от прокариот, то бактерий и архей нельзя объединять в одну группу) или эукариот (если признать прокариотами бактерий с археями, то эукариоты – потомки прокариот – получаются частью прокариот, что как бы странно); а учитывая, что эукариоты с большой вероятностью – смесь архей с бактериями, то каша получается совсем знатная. Не уверен, что читатель на этом месте не запутался, но систематика всегда была лабиринтом. Как часто бывает, при столкновении отличной идеи с реальностью приходится юлить и изобретать костыли.