
Палеонтология антрополога. Книга 2. Мезозой
В середине периода на первый план вышли другие оригинальные темноспондилы – Plagiosauroidea, например, Plagiosternum. Они имели короткую, зато крайне широкую – в 2,5 раза больше длины – голову с огромнейшими глазницами; видимо, это была защита от проглатывания капитозавроидами. А гарантией служил спинной панцирь из рельефных костных пластинок.

Aphaneramma

Mastodonsaurus
В конце триаса на обитателей озёр наводили страх последние гигантские темноспондилы Capitosauroidea (или Capitosauria). Paracyclotosaurus достигал 2,3 м и имел голову 60 см длиной, Metoposaurus и Jammerbergia formops – 2,5–3 м и 50–60 см, Anaschisma (она же Koskinonodon, Buettneria и Borborophagus) – 3 м и 65 см, а Mastodonsaurus – 4–6 м и 1,25 м! Это уже и для динозавра не зазорно. Такие огромные плюхи лежали на дне и подстерегали проплывающую мимо рыбу. Короткий хвост мог бросить тяжёлого хищника не некоторое расстояние, но не позволял долго гнаться за добычей. Коротенькие лапки уже не были способны двигать по суше тело весом в 450 кг. Ключицы и межключица разрастались в огромный ромбический щит, закрывавший всю грудь – почти как у черепахи, только снизу и лишь на половину туловища. Треугольные плоские головы с мелкими зубами и направленными вверх глазками работали как захлопывающиеся капканы, так же как у их предков эриопоидов.
В раннем триасе появляются более-менее узнаваемые пращуры лягушек. В оленекском веке по Мадагаскару ползали Triadobatrachus massinoti, а по Польше – Czatkobatrachus polonicus. Именно ползали, так как эти лягушки имели длинное тело и короткие ножки; у них ещё не срослись кости предплечья и голени, вполне сохранялись рёбра и свободные хвостовые позвонки, зато наружного хвоста уже не было (или ещё был, но уже совсем рудиментарный), и голова была уже почти как у современных квакушек. В норийском веке Аризоны тоже жили какие-то безымянные лягушки.
Как вы там, потомки?
Современные бесхвостые амфибии Anura – лягушки, жабы и квакши – весьма специализированы. Их локтевая кость срослась с лучевовй, а большая берцовая – с малой, хвостовые позвонки слились в единый уростиль, туловище укорочено, а рёбер вовсе нет. Подвздошная кость таза подвижна и работает как дополнительный рычаг для пущего прыгания, а многие к тому же умеют выбрасывать язык при охоте. В триасе не было ничего из этого, но всё же базовый план строения узнаётся именно как лягушачий.

Сухопутная фауна рептилий на границе эр резко сократилась. Из былого великолепия остались почти исключительно мелкие насекомоядные формы, а крупные – только текодонты Proterosuchidae и дицинодонты Lystrosauridae. Впрочем, в отсутствие конкуренции разнообразие быстро восстанавливалось.
Продолжали своё бытие парарептилии Procolophonia, например, Procolophon – короткая широкая коротконогая и короткохвостая «ящерица» с треугольной глазастой головой с длинными отростками на скулах. У Hypsognathus fenneri и Leptopleuron lacertinum скулы покрылись уже острыми шипами. Проще выглядела Tichvinskia – без лишних украшений, зато с глазницами в две трети черепа (через которые заодно проходили жевательные мышцы) и огромным теменным глазом; от вида T. vjatkensis сохранился целый скелет, уютно свернувшийся в ископаемой норке. Проколофоны любопытны своими зубами: передние были колышкообразными, а задние – расширенными поперечно и двувершинными. Некоторые проколофоны, например, Procolophon laticeps, Sauropareion anoplus, Owenetta kitchingorum и Candelaria barbouri независимо обрёли височные окна, хотя формально парарептилии относятся к анапсидам, то есть рептилиям без височных окон.
В триасе из каких-то анапсидных парарептилий возникли черепахи Testudines (или Chelonia).
Маленькая тонкость
На тему происхождения черепах написаны десятки статей и книг, но до обнаружения строго промежуточных форм многое оставалось неясным. Проблема в том, что современные черепахи крайне специализированы, кости их черепа очень рано срастаются без всяких видимых швов, остальной скелет тоже весьма своеобразен. Чего стоит только строение панциря – прирастание фактически окостеневшей кожи к позвоночнику и рёбрам.
Кстати, из-за панциря черепахи потеряли рёберное дыхание и вынуждены дышать горлом, которое у них не такое уж большое. Но крайне низкий обмен веществ спасает положение – много кислорода черепахам и не надо. Зато можно задерживать дыхание на полчаса, впадать в спячку, не есть неделями и жить сотни лет.
Некоторые черепахи пошли ещё дальше. Китайский трионикс Pelodiscus sinensis имеет в ротовой полости тонкие ворсинки, работающие одновременно как жабры и как средство выведения из организма мочевины. С таким приспособлением можно сидеть в воде и по 15 часов безвылазно! Трионикс Apalone spinifera, черепаха Фитцроя Rheodytes leukops и некоторые другие освоили дыхание обратной частью тела – стенками клоаки. Для лучшей эффективности в клоаке даже могут возникать специальные расширения – клоакальные сумки, поверхность которых покрыта ветвящимися сосочками, богатыми капиллярами. Чем не лёгкие? Суммарно подводное дыхание составляет для взрослых черепах Фитцроя 40 %, а для детишек – аж 73 %! В общем-то, это больше, чем у многих рыб. Так что черепахи доказали – потеря жабр не необратима.
И как можно после таких достижений называть черепах примитивными!..
Великий предок всех черепах – германский Pappochelys rosinae из ладинского века (около 240 млн л. н.). Он ещё был очень похож на тяжеловесную ящерицу с коротким широким туловищем, длинной шеей и очень длинным хвостом, у него было много зубов, но не было панциря. Черепаху в паппохелисе – «черепашьем дедушке» в переводе с греческого – можно узнать лишь по тонким деталям строения, из коих самый наглядный – крайне расширенные рёбра. Эволюционную эстафету перенял в целом очень похожий китайский Eorhynchochelys sinensis из начала карнийского века (228 млн л. н.), который тоже ещё не обрёл панцирь, зато успел потерять все зубы в передней части челюстей, где их заменил клюв; впрочем в задней части челюстей зубы ещё сохранялись. Ещё немножко – и китайский Odontochelys semitestacea 220 млн л. н., сохраняя зубы, обрёл панцирь, но, как ни странно, только нижний, то есть пластрон, тогда как верхнего – карапакса – ещё не было. Скелеты одонтохелисов найдены в морских отложениях; то ли они плавали по поверхности и им было актуальнее защищать пузико от посягательств снизу, то ли спинной панцирь на самом деле имелся, но только кожаный, как у мягкотелых черепах Trionychoidea и кожистых черепах Dermochelyidae. Некоторые учёные даже предполагают, что панцирь у одонтохелиса уже успел исчезнуть для пущей прыткости из-за жизни в море.
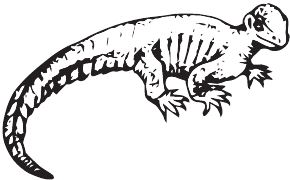
Pappochelys rosinae
Наконец, германский Proganochelys quenstedti (он же Psammochelys, Stegochelys и Triassochelys) 210 млн л. н. имел уже хорошо развитые верхний и нижний панцири, соединённые швом, причём карапакс срастался с остистыми отростками позвонков и тазом, а сзади был украшен торчащими назад шипами. Голова и хвост тоже были дополнительно защищены шипами, на нёбе ещё сохранялись маленькие зубки. Проганохелисы были немаленькими – практически квадратный панцирь имел длину и ширину до 65 см. В отличие от плавающего одонтохелиса, проганохелис был наземным животным. Примерно в то же время – в норийском веке – и тоже в Германии и Польше жили в целом похожие Proterochersis (они же Murrhardtia) и Keuperotesta limendorsa, а в Аргентине – Palaeochersis talampayensis, только они были поменьше – 30–45 см, их овальные выпуклые панцири не имели никаких украшений, а зубы окончательно пропали.
Маленькая тонкость
Черепахи – отличный пример для расчёта скорости эволюции. 240 миллионов лет назад паппохелис ещё был совсем не похож на черепаху, 230–220 млн л. н. эуринхохелис и одонтохелис начали обзаводиться панцирем и терять зубы, а 210 млн л. н. проганохелис и его родственники – уже образцовые черепахи. То есть, понадобилось каких-то 30 миллионов лет для полного преображения «под ключ». Показательно, что оценки для многих других эволюционных свершений вполне сопоставимы – возникновение амфибий, китов и даже людей (от совсем-совсем обезьяны до совсем-совсем человека) тоже укладывается в сроки в 20–30 миллионов лет.
Как сплошь и рядом случалось в эволюции, черепахи, возникнув, достигли своей черепашьей нирваны и в последующем менялись совсем незначительно. Конечно, всякие полезные мелочи (например, водное дыхание или подвижный сустав двух половин пластрона у шарнирных черепах Сuorа) добавлялись, но общий принцип оставался прежним: туповатая коробочка на кривых коротких ножках. В итоге черепахи, хотя часто называются самыми примитивными современными рептилиями, удивительным образом сочетают архаичность в одних чертах и резкую специализацированность, то есть прогрессивность, в других.
Развесистый куст возможностей реализовали диапсиды, давшие множество специфических групп. У части височные окна так сильно преобразились (нижнее закрылось), что иногда их выделяют в отдельную группу парапсид, или эвриапсид.
Опустевшее море в раннем триасе освоили Ichthyopterygia. Конечно, там ещё водились акулы, но, во‐первых, далеко не в том количестве, что раньше, а во‐вторых, вкусных фолидофорусов и лептолеписов хватало на всех. Первые ихтиоптеригии – Hupehsuchia из раннего (с оленекского века) и среднего триаса Китая, например, Hupehsuchus nanchangensis, Eohupehsuchus brevicollis и Nanchangosaurus suni, как водится, были похожи на длинно-остромордых крокодилов разной степени вытянутости и толщины с бронированной спиной, огромными широкими кистями и стопами, использовавшимися как вёсла, и мощным гребным хвостом. У многих хупехзухий увеличивался предпервый палец, а один из скелетов оказался обладателем семи пальцев на передней лапе и шести – на задней. Вероятно, такие пальцы уже не были самостоятельны, а образовывали широкие ласты. Рёбра Parahupehsuchus longus были расширены, плотно уложены и покрыты сплошным панцирем из мелких окостенений. Спина Eretmorhipis carrolldongi тоже была покрыта здоровенными костными бляхами, а голова оказалась удивительно похожей на голову утконоса: верхняя челюсть расширена и раздвоена, а между половинками залегала дополнительная косточка, аналогичная «парадоксальной косточке» (os paradoxum) утконоса.

Eretmorhipis carrolldongi
Хупехзухии оказались, хотя и оригинальными, но эндемичными животными, не давшими развития. Самой же успешной группой триасовых ихтиоптеригий стали ихтиозавры Ichthyosauria (Ichthyosauromorpha, или Ichthyosauriformes).
Первые их представители из оленекского века (248 млн л. н.) Китая – Cartorhynchus lenticarpus и Sclerocormus parviceps – были ещё очень непохожи на тех «дельфинчиков», которых все привыкли видеть на буриановских картинах. Это были толстые неуклюжие создания с короткой широкой мордой, огромными глазами, широкими лапами-плавниками и длинным веслообразным хвостом. Спинного плавника они, по всей видимости, пока не обрёли. Возможно, они ещё умели выползать на берег, как тюлени.
Но очень быстро на основе таких неуклюжих тварей возникли настоящие покорители океана. Уже в следующем – анизийском – ярусе Японии и Канады обнаруживается Utatsusaurus hattai. Этот трёхметровый ихтиозавр всё ещё был широкоголов и не имел спинного плавника, но обладал всеми узнаваемыми чертами группы: вытянутым рылом, зачаточным хвостовым плавником, причём позвоночный столб ушёл в нижнюю лопасть, а верхняя образована только кожей. Таз начал терять соединение с крестцом, а передние и задние ласты были почти равной длины, выдавая не очень прыткого пловца.

Cartorhynchus lenticarpus
Работа над ошибками
На реконструкциях ихтиозавры сплошь и рядом изображены как крутоспинные дельфинчики, бодро прыгающие и кувыркающиеся над пенистыми волнами. Очень эффектно, жаль только, что совершенно не реалистично. Позвоночник ихтиозавров, как и у всех рептилий, изгибался с боку на бок, а не сверху-вниз, так что они плавали подобно морским змеям, но никак не дельфинам. Именно поэтому и хвостовой плавник ихтиозавров ориентирован в вертикальной плоскости, как у рыб, а не горизонтальной, как у китообразных (конечно, и акулы с марлинами иногда сигают над волной, вертикальный хвост им не помеха, но всё же до дельфинчиков им далеко). К тому же рёбра ихтиозавров были длинными и сильно изогнутыми, стало быть, тело походило на бочку, передние и задние плавники же сплошь и рядом одинаковой длины. Всё это выдаёт не резвых пловцов, а надувные матрасы. В лучшем случае ихтиозавры могли «летать» под водой, как современные морские черепахи. Конечно, знатоки и ценители ихтиозавров обижаются на такой образ их любимых животных, но что поделать – с точки зрения антрополога, привыкшего к скачущим по ветвям приматам, бодрость морских рептилий совсем не впечатляет.
С ихтиозаврами в очередной раз сработал принцип «первого парня на деревне» – раньше всех застолбив свою экологическую нишу и в отсутствие достойных конкурентов, эти хищники быстро достигли совершенства по своим понятиям и до середины мелового периода принципиально уже не менялись. Недоработки конструкции аукнулись гораздо позже и стали очевидны лишь с появлением гораздо более совершенных морских ящеров. Но до этого было ещё очень далеко.
В последующем ихтиозавры стремительно расселяются по всему миру: раннетриасовый Chaohusaurus geishanensis из Китая, среднетриасовые Mixosaurus, жившие от Европы до Северной Америки и Индонезии, длиннейшие – до 10–12 м – Cymbospondylus и многие прочие начали бороздить воды морей. Название Himalayasaurus tibetensis говорит само за себя, очевидно, что география с триасовых времён сильно поменялась. Ясно, началась эпоха специализации. Весьма примитивная Grippia longirostris со Шпицбергена имела в передней части челюстей острые, а в задней – полушаровидные давящие зубы в два ряда, у метровых Phalarodon передние зубы были округлые приострённые, а задние мало отличались от моляров млекопитающих; такими было очень удобно разгрызать раковины моллюсков и панцири крабов. Десятиметровый Guanlingsaurus liangae (он же Shastasaurus liangae) и его родственники вообще потеряли все зубы. Вероятно, они всасывали планктон, широко разевая рот, наподобие усатых китов.
Плавники ихтиозавров до середины триаса ещё сохраняли узнаваемые пять пальцев, но плотно прижатых друг к другу в единой ласте, а число фаланг стало произвольным. У некоторых среднетриасовых ихтиозавров появился характерный треугольный спиной плавник, подобный акульему.
К концу триаса ихтиозавры достигли впечатляющих размеров: Shonisaurus popularis вырастал до 15 м, а похожий на него Shastasaurus sikkaniensis (он же Shonisaurus sikanniensis) – до 21 м! В их плавниках число пальцев сократилось до трёх, а количество фаланг резко выросло и стало совсем уж неопределённо большим. Эти гигантские поплавки неспешно рассекали воды и ловили рыбу своими вытянутыми острыми беззубыми челюстями, как пинцетами.
В среднем и позднем триасе к ихтиозаврам добавились гораздо более шустрые Thalattosauria. Опять крокодилообразные рептилии нырнули в воду: длинные тонкие, очень гибкие и ловкие пловцы с узкой вытянутой мордой, длинной шеей, умеренными лапами и мощным гребным хвостом до 60 позвонков. Впрочем, некоторые детали могли разниться. Например, двухметровый Thalattosaurus имел загнутую на конце морду, острые зубы спереди и плоские зубообразные выросты челюсти сзади; он, видимо, питался моллюсками. Остромордый Askeptosaurus italicus, судя по тонким зубам, ловил рыбу. Поздний Endennasaurus acutirostris вовсе потерял зубы, хотя строение остального скелета у него удивительно примитивное. Gunakadeit joseeae замечателен тем, что найден на Аляске, а судя по развитому подъязычному аппарату, имел длинный сильный язык, что наводит на мысли о каком-то экзотическом варианте питания. В целом похож Miodentosaurus brevis, только он был короткомордый и четырёхметровый. У Hescheleria rubeli передний конец короткой морды загибался совсем уж вертикально вниз, а зубов осталось совсем мало, зато на нижней челюсти торчал здоровенный треугольный «клык». А вот Xinpusaurus отличался гораздо сильнее: очень короткая шея, странно искривлённые челюсти, причём у вида X. kohi острая верхняя челюсть длиннее короткой нижней, с подобием хвостового плавника на кончике длиннейшего хвоста, он был больше похож на ихтиозавра.
Маленькая тонкость
Между прочим, хотя талаттозавры исходно диапсидные, то есть с двумя окнами в черепе, поздние представители потеряли верхнее окно. Это показывает возможности эволюции черепа в разные стороны и подкидывает дровишек в стародавний спор о первичности или вторичности анапсидности черепа черепах и их родственников. Может, и черепахи когда-то имели височные окна?
Многие триасовые водные рептилии настолько странны, что с трудом находят свою родню. Например, крайне странно выглядели двухметровый Helveticosaurus zollingeri и шестнадцатисантиметровый Eusaurosphargis dalassoi. У гельветикозавра была короткая треугольная голова на короткой шее, огромнейшие зубищи-сабли, длинное, но широкое тело, мощные лапы с увеличенным числом фаланг и длинный веслообразный хвост. Эузаросфаргис имел короткие притупленные зубы, странно не смыкающиеся челюсти (зубной ряд верхней задирался вверх, а нижняя челюсть была прямой), овальное тело с длинными тонкими рёбрами и крупночешуйчатым панцирем, а также короткий хвост, передняя часть которого была очень сильно расширена поперечно, а задняя торчала тоненькой ниточкой. Sinosaurosphargis yunguiensis был внешне почти неотличим от черепахи из-за своего круглого панциря, набранного из маленьких окостенений. Спинные и брюшные рёбра были расширены и сами по себе формировали почти сплошные верхний и нижний щиты. Однако его рот был усеян многочисленными мелкими зубками, на короткой плоской треугольной голове были огромные нижние височные окна, почти полностью закрытые верхние и отверстие для третьего глаза – совершенно нетипичное для черепах сочетание.
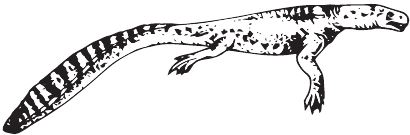
Helveticosaurus zollingeri
Ещё одна водная триасовая группа парапсид – Synaptosauria – включает, на первый взгляд, весьма разнородных существ. В чём-то похожа на хупехзухий китайская завроптеригия Atopodentatus unicus из анизийского века – очень длинная тонкая трёхметровая «дракошка» с необычайно расширенным «утиным» клювом и мелкими тупыми зубами, уложенными в плотную гребёнку, направленную строго вперёд. Судя по всему, атоподентатус питался морскими водорослями, которые старательно соскребал с камней своей зубной щёткой.
Работа над ошибками
Первая реконструкция атоподентатуса предполагала, что его верхняя челюсть была загнута под прямым углом вниз и при этом расщеплена надвое, а на обеих половинках располагались мелкие зубки, направленные друг к другу. Такого чуда никто не ожидал даже от китайских триасовых тварей! Предполагалось, что такая удивительная форма челюстей работала как фильтровальный аппарат, наподобие китового уса. Впрочем, сенсация продержалась не очень долго, всего пару лет. Новые находки показали, что первая реконструкция была основана на смещённых костях. В реальности морда атоподентатуса была скорее похожа на гипертрофированный утиный клюв.
Другой крайности специализации достигли Placodontia. Плакодонты освоили нишу медленных придонных поедателей моллюсков и брахиопод, болтавшихся в прибойной зоне морей. Более примитивные двухметровые Placodus были тяжеловесными коротконогими короткошеими и длиннохвостыми животными, спина их была покрыта костными пластинками. Конечно, самое удивительное в их строении – зубная система. На широкой морде плакодуса спереди торчали редкие шпателевидные резцы, по бокам челюстей тянулись ряды по четыре овальных плоских давящих зуба, а на нёбе с каждой стороны располагались по три огромных зуба-жернова. Очевидно, передними долотами плакодусы отскребали раковины с камней, а задними давили их. То-то хруст стоял при этом!
Впечатляет грудная клетка Paraplacodus broilii: к длинным поперечным отросткам позвонков крепились расширенные рёбра, шедшие сначала строго вбок, а потом под прямым углом поворачивавшие вниз; снизу коробку замыкали так же изогнутые брюшные рёбра, так что вся конструкция имела вид коробки-параллелепипеда.
Маленькая тонкость
Как бы странно ни выглядели плакодонты, и для них находятся аналоги в длинной череде странных животных. Кайнозойские млекопитающие Palaeoparadoxia, Desmostylus и их родственники тоже были медлительными прибрежными жевателями моллюсков с огромными стамесками в передней части челюстей и бугристыми давилками в задней.
Более продвинутые плакодонты пошли ещё дальше, запаковавшись в панцири. У среднетриасового Cyamodus и позднетриасовой Psephoderma большая часть тела была закрыта почти круглым панцирем из плотно уложенных костяных бляшек, а зад был прикрыт дополнительным широким щитом меньшего размера; очень длинный тонкий хвост тоже был бронирован. Головы этих странных существ имели форму луковицы за счёт огромных скуловых дуг и тонкого вытянутого носа, который в сочетании с круглыми направленными вперёд глазами придавал зверушкам ехидно-клоунское выражение. Нёбо было оборудовано парой огромных зубов-давилок и парой маленьких, а края челюстей – молотилками калибром поменьше. Голова Placochelys placodonta была в целом похожа на головы предыдущих тварей, только с роговым клювом спереди вместо резцов и с короткими шипами по заднему краю, а пятиугольный панцирь был уже единым, хотя по-прежнему набранным из отдельных многоугольных щитков, среди которых выделялись более крупные пирамидки. Хвост такой псевдочерепахи укоротился, а длинные пальцы на лапах с большой вероятностью объединились в сплошные овальные ласты.
Апофеозом явился позднетриасовый германский Henodus chelyops. У него сохранилась лишь пара задних зубов на верхней челюсти и пара – на нижней, а три края строго прямоугольной головы оказались обрамлены острым роговым клювом. Плоский округло-квадратный панцирь закрывал всё тело, кроме шеи с головой и довольно длинного хвоста, для которых спереди и сзади на панцире были предусмотрены глубокие вырезки. Щит был собран из мозаики костяных пластинок с роговым покрытием.

Henodus chelyops
Как видим, медлительность и уязвимость быстро вызвали необходимость бронирования. С другой стороны, существование черепахоподобных животных однозначно указывает на наличие опасных злобных хищников; возможно, таким проклятием плакодонтов были саблезубые гельветикозавры.
Большая часть синаптозавров – Sauropterygia. В их числе Nothosauria – морские рептилии довольно банального облика: вытянутое тело, длинные шея и хвост, узкая плоская вытянутая голова с длинными острыми зубами, иногда далеко перекрещивающимися друг с другом. Судя по умеренной длины лапам с развитыми пальцами, нотозавры вполне могли выползать на берег; впрочем, у некоторых кисти и стопы могли сливаться в сплошные ласты. Нотозавры были одними из самых обычных животных морских побережий, иногда их скелеты находят лежащими чуть ли не слоями друг на друге. Nothosaurus вырастал от 3,5 до 7 м, Ceresiosaurus – от 1 до 3 м, Lariosaurus – до 60 см, а некоторые Neusticosaurus (они же Pachypleurosaurus) – всего до 18 см.
Среднетриасовый Pistosaurus longaevus и в особенности Bobosaurus forojuliensis из начала карнийского века Италии – переходные звенья между нотозаврами и плезиозаврами, их лапы окончательно стали ластами, а тело укоротилось и потеряло гибкость. С их появлением забрезжила эпоха новых морских царей – плезиозавров.