
Life on Earth

Near horizontal layers of sedimentary rock, cut through by the Colorado River, forming the Grand Canyon, USA.
The deepest cleft that exists in the earth’s surface is the Grand Canyon in the western United States. The rocks through which the Colorado River has cut its way still lie roughly horizontally, layer upon layer, red, brown and yellow, sometimes pink in early light, sometimes blue in the shadowed distance. The land is so dry that only isolated juniper trees and low scrub freckle the surface of the cliffs, and the rock strata, some soft, some hard, are clear and stark. Most of them are sandstones or limestones that were laid down at the bottom of the shallow seas that once covered this part of North America. When they are examined closely, breaks in the succession can be detected. These represent times when the land rose, the seas drained away and the seabed became dry so that the deposits that had accumulated on it were eroded away. Subsequently, the land sank again, seas flooded back and deposition restarted. In spite of these gaps, the broad lines of the fossil story remain clear.
A mule will carry you in an easy day’s ride from the rim to the very bottom of the Canyon. The first rocks you pass are already some 200 million years old. There are no remains of mammals or birds in them, but there are traces of reptiles. Close by the side of the trail, you can see a line of tracks crossing the face of a sandstone boulder. They were made by a small four-footed creature, almost certainly a lizard-like reptile, running across a beach. Other rocks, at the same level elsewhere, contain impressions of fern leaves and the wings of insects.
Halfway down the Canyon, you come to 400-million-year-old limestones. There are no signs of reptiles to be found here, but there are the bones of strange armoured fish. An hour or so later – and a hundred million years earlier – the rocks contain no sign of backboned animals of any kind. There are a few shells and worms that have left behind a tracery of trails in what was the muddy seafloor. Three-quarters of the way down, you are still descending through layers of limestone, but now there is no sign of fossilised life whatever. By the late afternoon, you ride at last into the lower gorge where the Colorado River runs green between high rock walls. You are now well over a vertical kilometre below the rim, and the surrounding rocks have been dated to the immense age of 2,000 million years. Here you might hope to find evidence for the very beginnings of life. But there are no organic remains of any kind. The dark fine-grained rocks lie not in horizontal layers like all those above, but are twisted and buckled and riven with veins of pink granite.
Are signs of life absent because these rocks and the limestones directly above are so extremely ancient that all such traces have been crushed from them? Could it be that the first creatures to leave any sign of their existence were as complex as worms and molluscs? For many years these questions puzzled geologists. All over the world, rocks of this antiquity were carefully searched for organic remains. One or two odd shapes were found, but most authorities dismissed these as patterns produced by the physical processes of rock formation that had nothing whatever to do with living organisms. Then during the 1950s, the searchers began to use high-powered microscopes on some particularly enigmatic rocks.
Around 1,600 kilometres northeast of the Grand Canyon, ancient rocks of about the same age as those beside the Colorado River outcrop on the shores of Lake Superior. Some of them contain seams of a fine-grained flint-like substance called chert. This was well known during the nineteenth century because the pioneers used it in their flintlock guns. Here and there, it contains strange white concentric rings a metre or so across. Were these merely eddies in the mud on the bottom of the primeval seas, or could they have been formed by living organisms? No one could be sure and the shapes were given the noncommittal name of stromatolite, a word derived from Greek meaning no more than ‘stony carpet’. But when researchers cut sections of these rings, ground them down into slices so thin that they were translucent and examined them through the microscope, they found, preserved in the chert, the shapes of simple organisms, each no more than one or two hundredths of a millimetre across. Some resembled filaments of algae; others, while they were unmistakably organic, had no parallels with living organisms; and some looked to be identical with the simplest form of life existing today: bacteria.
It seemed almost impossible to many people that such tiny things as microorganisms could have been fossilised at all. That relics of them should have survived for such a vast period of time seemed even more difficult to believe. The solution of silica which had saturated the dead organisms and solidified into chert was clearly as fine-grained and durable a preservative as exists. The discovery of the fossils in the Gunflint Chert stimulated further searches not only in North America but all over the world, and other microfossils were found in cherts in Africa and Australia. Some of these, astonishingly, pre-dated the Gunflint specimens by a billion years, and some scientists now claim to have found fossils from around 4 billion years ago, not long after the formation of the earth. But if we want to consider how life arose, fossils cannot help us, for the origin of life involved the interaction of molecules, which leave no fossil traces. To understand what scientists think happened we have to look back beyond even the earliest microfossils, to a time when the earth was completely lifeless.
In many ways the planet then was radically different from the one we live on today. There were seas, but the way the land masses lay bore no resemblance in either form or distribution to modern continents. Volcanoes were abundant, spewing noxious gases, ash and lava. The atmosphere consisted of swirling clouds of hydrogen, carbon monoxide, ammonia and methane. There was little or no oxygen. This unbreathable mixture allowed ultraviolet rays from the sun to bathe the earth’s surface with an intensity that would be lethal to modern animal life. Electrical storms raged in the clouds, bombarding the land and the sea with lightning.
Laboratory experiments were made in the 1950s to discover what might happen to these particular chemical constituents under such conditions. Such gases, mixed with water vapour, were subjected to electrical discharge and ultraviolet light. After only a week of this treatment complex molecules were found to have formed in the mixture, including sugars, nucleic acids and amino acids, the building blocks of proteins. We now know that such simple organic molecules can be found throughout the universe, including on interstellar bodies such as comets. But amino acids are not life, nor are they even necessary for life to exist. The experiment proved little about the origin of life.
All forms of life that exist today share a common way of transmitting genetic information, of telling cells what to do. It is a molecule called deoxyribonucleic acid, or DNA for short. Its structure gives it two key properties. First, it can act as a blueprint for the manufacture of amino acids; and second, it has the ability to replicate itself. With this substance, molecules had reached the threshold of something quite new. These two characteristics of DNA also characterise even the simplest of living organisms such as bacteria. And bacteria, besides being the simplest form of life we know, are also among the oldest fossils we have discovered.
The ability of DNA to replicate itself is a consequence of its unique structure. It is shaped like two intertwined helices. During cell division, these unzip, splitting the molecule along its length into two separate helices. Each then acts as a template to which other simpler molecules become attached until each has once more become a double helix.
The simple molecules from which the DNA is mainly built are of only four kinds, but they are grouped in trios and arranged in a particular and significant order on the immensely long DNA molecule. This order specifies how the twenty or so different amino acids are arranged in a protein, how much is to be made, in what tissue and when. A length of DNA bearing such information for a protein, or for how a protein should be expressed, is called a gene.
Occasionally, the DNA copying process involved in reproduction may go wrong. A mistake may be made at a single point, or a length of DNA may become temporarily dislocated and be reinserted in the wrong place. The copy is then imperfect and the proteins it will create may be entirely different. Changes in the DNA sequence can also be induced by chemicals or radiation. When this occurred in the first organisms on earth, evolution began, for such hereditary changes, brought about by mutation and errors, are the source of variations from which natural selection can produce evolutionary change.
Because all life shares DNA as the hereditary material, it is possible to compare DNA sequences in different organisms and show how they are related. Such is the progress of technology that it is now also possible to sequence all the DNA in an organism in a matter of hours, using a device the size of a mobile phone. The millions of DNA sequences that have been established, stored in databases and compared show us unequivocally that, just as Darwin predicted, all life on earth shares a common ancestor. Because parts of our DNA accumulate mutations at a constant rate, like a molecular clock, we can use DNA sequences to estimate when two species split apart. In general, genetic and fossil timings agree with each other, although genetic data do sometimes throw up surprises. Using this method we can estimate that the Last Universal Common Ancestor of all life on earth – commonly known as LUCA, and basically a population of simple bacteria – lived around 4 billion years ago. Everything we can see around us can trace its ancestry back to that group of cells.
Such vast periods of time baffle the imagination, but we can form some idea of the relative duration of the major phases of the history of life if we compare the entire span, from these first beginnings until today, with one year. That means that, roughly, each day represents around ten million years. On such a calendar, the Gunflint fossils of algae-like organisms, which seemed so extremely ancient when they were first discovered, are seen to be quite late-comers in the history of life, not appearing until the second week of August. In the Grand Canyon, the oldest worm trails were burrowed through the mud in the second week of November and the first fish appeared in the limestone seas a week later. The little lizard will have scuttled across the beach during the middle of December and humans did not appear until the evening of 31 December.
But we must return to January. The bacteria fed initially on the various carbon compounds that had taken so many millions of years to accumulate in the primordial seas, producing methane as a by-product. Similar bacteria still exist today, all over the planet. And that was all there was, for around five or six months of our year. Then, in the early summer of the year of life, so some time over 2 billion years ago, bacteria developed an amazing biochemical trick. Instead of taking ready-made food from their surroundings, they began to manufacture their own within their cell walls, drawing the energy needed to do so from the sun. This process is called photosynthesis. One of the ingredients required by the earliest form of photosynthesis is hydrogen, a gas that is produced in great quantities during volcanic eruptions.
Conditions very similar to those in which the early photosynthesising bacteria lived can be found today in such volcanic areas as Yellowstone in Wyoming. Here a great mass of molten rock, lying only a thousand metres or so, down in the earth’s crust, heats the rocks on the surface. In places, the ground water is well above boiling point. It rises up channels through the rocks under decreasing pressure until suddenly it flashes into steam and water spouts high into the air as a geyser. Elsewhere, the water wells up into steaming pools. As it trickles away and cools, the salts it gathered from the rocks on its way up, together with those derived from the molten mass far below, are deposited to form rimmed and buttressed basins, surrounded by tiers of terraces. In these scalding mineral-laden waters, bacteria flourish. Some grow into matted filaments and curds, others into thick leathery sheets. Many are brilliantly coloured, their intensity of hue varying during the year as the colonies wax and wane. The names given to these pools hint at the variety of the bacteria and the splendour of the effects they produce – Emerald Pool, Sulphur Cauldron, Beryl Spring, Firehole Falls, Morning Glory Pool and – a particularly rich one with several species of bacteria – Artists’ Paintpots.

Hot spring, its water coloured by bacteria, Yellowstone National Park, Wyoming, USA.
When you wander through this amazing landscape, you can smell sulphurated hydrogen, the unmistakable stench of rotting eggs, produced by the reaction of ground water with the molten rock far beneath. This is the source from which many of the bacteria here obtain their hydrogen, and as long as bacteria were dependent upon volcanic action for it, they could not spread widely. But other forms eventually arose which were able to extract hydrogen from a very much more widespread source – water. This development was to have a profound effect on all life to come, for if hydrogen is extracted from water, the element that remains is oxygen. The organisms that did this are barely more complex in structure than bacteria. They are sometimes called blue-green algae because they appeared to be close relatives of the green algae that are common in ponds, but now we realise they are similar to the ancestors of those algae, and they are referred to as cyanobacteria or, simply, blue-greens. The chemical agent which they contain, making it possible for them to use water in the photosynthetic process, is chlorophyll, which is also possessed by true algae and plants.
Blue-greens are found wherever there is constant moisture. You can often see mats of them, beaded with silver bubbles of oxygen, blanketing the bottoms of ponds. In Shark Bay, on the northwest coast of tropical Australia, they have developed in a particularly spectacular and significant form. Hamelin Pool, one small arm of this vast inlet, has its entrance blocked by a sand bar covered with eel grass. The flow of water in and out of the Pool is so greatly impeded that evaporation under the grilling sun has made the waters very salty indeed. As a result, marine creatures such as molluscs which would normally feed on blue-greens and keep them in check, cannot survive. The blue-greens, therefore, flourish uncropped just as they did when they were the most advanced form of life anywhere in the world. They secrete lime, forming stony cushions near the shores of the Pool and teetering columns at greater depths. Here is the explanation of those mysterious shapes seen in section in the Gunflint Chert. The blue-green pillars of Hamelin Pool are living stromatolites, and the groups of them standing on the sun-dappled seafloor are as close as we may ever get to a scene from the world of 2 billion years ago.
The arrival of the blue-greens marked a point of no return in the history of life. In ways we do not fully understand, the oxygen they produced eventually accumulated over the millennia to form the kind of oxygen-rich atmosphere that we know today. Our lives, and those of all other animals, depend on it. We need it not only to breathe but to protect us. Oxygen in the atmosphere forms a screen, the ozone layer, which cuts off most of the ultraviolet rays of the sun.
Life remained at this stage of development for a vast period. Then, around 2 billion years ago one single-celled life form found itself trapped inside another, in an entirely chance encounter. You can find examples of the kind of organisms it eventually produced in almost any patch of fresh water.
A drop from a pond, viewed through a microscope, swarms with tiny organisms, some spinning, some crawling, some whizzing across the field of vision like rockets. As a group they are often called the protozoa, or protists – the name means ‘first animals’, although they are now seen as a very disparate group, not all of which have any affinity with animals. They are all single cells, yet within their cell walls they contain much more complex structures than any bacterium possesses. One central packet, the nucleus, is full of DNA. This appears to be the organising force of the cell. Elongated bodies, the mitochondria, provide energy by burning oxygen in much the same way as many bacteria do. Many cells have a thrashing tail attached to them and this resembles a filamentous bacterium called a spirochete. Some also contain chloroplasts, packets of chlorophyll which, like blue-greens, use the energy of sunlight to assemble complex molecules as food for the cell. Each of these tiny organisms thus appears to be a committee of simpler ones. This, in effect, is what they are. The mitochondria are the descendants of the single-celled organism that was trapped some 2 billion years ago, say in June in the year of life, while the chloroplasts are descended from a trapped blue-green.
Protozoans reproduce by splitting into two, as bacteria do, but their internal structure is much more complex and their division, not surprisingly, is consequently an elaborate business. Most of the separate structures, the members of the committee, themselves split. Indeed, the mitochondria and chloroplasts, each with their own DNA as befits their origins as separate organisms, often do so independently of the division of the main cell. The DNA within the nucleus replicates in a particularly complex manner which ensures that all its genes are copied and that each daughter cell receives a complete duplicate set. There are, however, several other methods of reproduction practised by various protozoans on occasions. The details vary. The essential feature of all the techniques is that a shuffling of genes is involved. In some cases this takes place when two cells join up and exchange genes before breaking apart and then undergoing cell division some time later. In other cases, cells normally contain two complete sets of genes which, after shuffling, divide to make new cells with only one set. These cells are of two types – a large comparatively immobile one, and a smaller active one, driven by a flagellum. The first is called an egg and the second a sperm – for this is the dawning of sex. When the two types unite in a new amalgamated cell the genes are once again in two sets but in new combinations with genes from not just one parent but two. This may well be a unique combination which will produce a slightly different organism with new characteristics. Since the evolution of sex increased the possibilities of genetic variation, it also greatly accelerated the rate at which evolution could proceed as organisms encountered new environments.
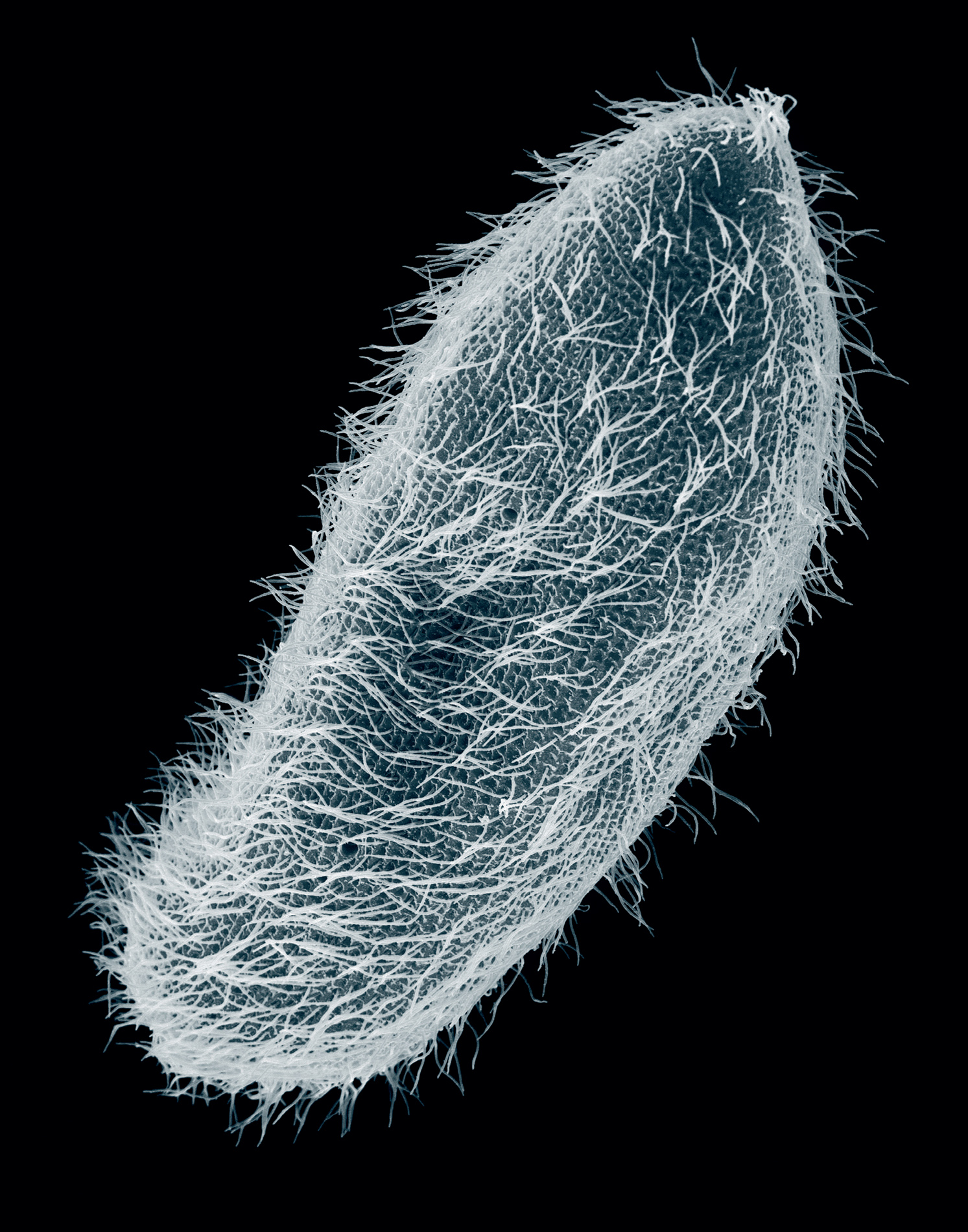
Ciliated protozoan (Paramecium multimicronucleatum), scanning electron micrograph (SEM).
There are tens of thousands of species of protozoans. Some are covered by a coat of flailing threads or cilia, which with a coordinated beat drive the creature through the water. Others, including the amoeba, move by bulging out fingers from the main body and then flowing into them. Many of those that live in the sea secrete shells with the most elaborate structure of silica or calcium carbonate. These are among the most exquisite objects that the microscope-carrying explorer will ever encounter. Some resemble minuscule snail shells, some ornate vases and bottles. The most delicate of all are of shining translucent silica, concentric spheres transfixed by needles, gothic helmets, rococo belfries and spiked space capsules. The inhabitants of these shells extend long threads through pores with which they trap particles of food.
Other protists feed in a different way, photosynthesising with the aid of their packets of chlorophyll. These can be regarded as plants; the remainder of the group, which feed on them, as animals. The distinction between the two at this level, however, does not have as much meaning as such labelling might suggest, for there are many species that can use both methods of feeding at different times.
Some protists are just large enough to see with the naked eye. With a little practice, the creeping grey speck of jelly which is an amoeba can be picked out in a drop of pond water. But there is a limit to the growth of a single-celled creature, for as size increases, the chemical processes inside the cell become difficult and inefficient. Size, however, can be achieved in a different way – by grouping cells together in an organised colony.
One species that has done this is volvox, a hollow sphere, almost the size of a pinhead, constructed from a large number of cells, each with a flagellum. The striking thing about these units is that they are virtually the same as other single cells that swim by themselves and have separate existences. The constituent cells of volvox, however, are coordinated, for all the flagella around the sphere beat in an organised way and drive the tiny ball in a particular direction.
This kind of coordination between constituent cells in a colony was taken a stage further, probably between 800 and 1,000 million years ago – some time in October in our calendar – when sponges appeared. Sponges can grow to a very considerable size. Some species form soft shapeless lumps on the seafloor two metres or so across. Their surfaces are covered with tiny pores through which water is drawn into the body by flagella, and then expelled through larger vents. The sponge feeds by filtering particles from this stream of water passing through its body. The colonial bonds between its constituents are very loose. Individual cells may crawl about over the surface of the sponge like amoebae. If two sponges of the same species are growing close to one another, they may, as they grow, come into contact and eventually merge into one huge organism. If a sponge is forced through a fine gauze sieve so that it is broken down into separate cells, these will eventually reorganise themselves into a new sponge, each kind of cell finding its appropriate place within the body. Most remarkably of all, if you take two sponges of the same species and treat them both in this extreme way and then mix cells from the two, they will reconstitute themselves into a single mixed-parentage entity.

Massive barrel sponge (Xestospongia testudinaria) and diver. Tubbataha Reef National Marine Park, Palawan, Philippines.
Some sponges produce a soft, flexible substance around their cells which supports the whole organism. This, when the cells themselves have been killed by boiling and washed away, is what we use in our baths. Other sponges secrete tiny needles, called spicules, either of calcium carbonate or silica, which mesh together to form a scaffold in which the cells are set. How one cell orientates itself and produces its spicule so that it fits perfectly into the overall design is totally unknown. When you look at a complex sponge skeleton such as that made of silica spicules which is known as Venus’ flower basket, the imagination is baffled. How could quasi-independent microscopic cells collaborate to secrete a million glassy splinters and construct such an intricate and beautiful lattice? We do not know. But even though sponges can produce such miraculous complexities as this, they are not like other animals. They have no nervous system, no muscle fibres. The simplest creatures to possess these physical characteristics are the jellyfish and their relatives.