
The Tangled Tree: A Radical New History of Life
It was now February 1858. Hardly anyone at that point recognized Darwin for what he was—an evolutionary theorist, in secret—and though Lyell was among that small group who did, as a close friend and confidant, Alfred Wallace certainly wasn’t. Charles Darwin to him was just a conventionally eminent naturalist, author of the Beagle chronicle and other safe books, including several on the taxonomy of barnacles. But a Dutch mail boat would soon stop at the port of Ternate, in the Moluccas, where Wallace had fetched up. He was excited by his own discovery, if it was a discovery, and eager to share this dangerous hypothesis with the scientific world. So he packed up his paper with a cover letter and mailed the packet to Mr. Darwin, hoping that Darwin might find it worthy. If so, maybe Darwin would share it with Mr. Lyell, who might help get it published.
The packet reached Darwin, probably on June 18, 1858, and hit him like a galloping ox. He felt crushed, scooped, ruined—but also honor-bound to grant Wallace’s request, passing the paper on toward publication. It would mean, Darwin knew, letting the younger man take all the credit for this epochal idea he himself had incubated for twenty years but was not yet quite ready to publish. Despite that, he did send the Wallace paper along to Lyell—communicating yelps of his own anguish along with it. Lyell took not just the paper but also the hint. Along with another of Darwin’s close scientific allies, the botanist Joseph Hooker, Lyell talked Darwin back from despair, suggested a posture of sensible fairness rather than self-abnegating honor, and brokered a compromise of shared credit. The result was a clumsily conjoined presentation—a pastiche of Wallace’s paper plus excerpts from Darwin’s unpublished writings—before a British scientific club, the Linnean Society, in the summer of 1858. Lyell and Hooker offered an introductory note, and then simply watched and listened. Proxies read the works aloud, with neither of the authors present. (Darwin was at home, where his youngest son had just died of scarlet fever; Wallace was still out in the far boonies of the Malay Archipelago.) This joint presentation made almost no impression on anyone, not even the few dozen Linnean members in attendance, because the night was hot, the language was obscure, the logic was elliptical, and the big meaning didn’t jump forth.
Seventeen months later, Darwin published On the Origin of Species. That 1859 book, not the 1858 paper or excerpts, launched the Darwinian revolution. It was only an abridged and hasty abstract of the much longer (and more tedious) book on natural selection that Darwin had been writing for years, but The Origin was just enough, in the right form, at the right time. It presented the theory as “one long argument,” not just a bare syllogism, and with oodles of data but not many footnotes. It was plainspoken, and readable by any literate person. It became a bestseller and went into multiple editions. It converted a generation of scientists to the idea of evolution (though not to natural selection as the prime mechanism). It was translated and embraced in other countries, especially Germany. That’s why Darwin is still history’s most venerated biologist and Alfred Russel Wallace is a cherished underdog, famous for being eclipsed, to the relatively small subset of people who have heard of him.
The crux of the “one long argument” comes in chapter 4 of The Origin, titled “Natural Selection,” in which Darwin describes the central mechanism of his theory. It’s the same combination of three principles that he had scratched into his notebook two decades earlier, plus the turned crank. “Natural selection,” he wrote in the book, “leads to divergence of character and to much extinction of the less improved and intermediate forms of life.” Lineages change over time, he stated. You could see that in the fossil record. Different creatures adapt to different niches, different ways of life, and thereby diversify into distinct forms and behaviors. Transitional stages disappear. Then he wrote: “The affinities of all the beings of the same class have sometimes been represented by a great tree. I believe this simile largely speaks the truth.”
8
Darwin explored the tree simile in one extended paragraph, ending that chapter of The Origin. “The green and budding twigs may represent existing species,” he wrote. From there he worked backward: woody twigs and small branches as recently extinct forms; competition between branches for space and for light; big limbs dividing into branches, then those into lesser branches; all ascending and spreading from a single great trunk. “As buds give rise by growth to fresh buds,” Darwin wrote, and those buds grow to be twigs, and those twigs grow to be branches, some vigorous, some feeble, some thriving, some dying, “so by generation I believe it has been with the great Tree of Life, which fills with its dead and broken branches the crust of the earth, and covers the surface with its ever branching and beautiful ramifications.” There’s a nice word: ramifications.
It’s especially good in this context because, while the literal definition is “a structure formed of branches,” from the Latin ramus, of course the looser definition is “implications.” Darwin’s tree certainly had implications.
Furthermore his book, like Edward Hitchcock’s, included a treelike illustration. This was the only illustration, the only graphic image of any sort, in the first edition of The Origin. It appeared between pages 116 and 117, amid his discussion of how lineages diverge over time. A foldout, again like Hitchcock’s, but published in simple black and white. It was a schematic figure, not an artfully drawn tree, not even so lively as the little sketch in his notebook long ago. Darwin called it a diagram. It showed hypothetical lineages, proceeding upward through evolutionary time and diverging—that is, dotted lines, rising vertically and branching laterally. Darwin was no artist, but, even lacking such talent, he could have laid out this diagram with a pencil and a ruler. In its draft version, as sent to the lithographer, he probably had. But it made the arboreal point.
Each increment of vertical distance on the ruled page, Darwin explained, stood for a thousand generations of inheritance. Deep time. Eleven major lineages began the ascent. Eight of those came to dead ends—meaning, they went extinct. Trilobites, ammonites, ichthyosaurs, and plesiosaurs had all suffered such ends, leaving no descendants of any sort. One lineage rose through the eons without splitting, without tilting, like a beanstalk—meaning that it persisted through time, unchanged. That’s much the way horseshoe crabs, sometimes called living fossils, have survived relatively unchanged (at least externally, so far as fossilization can show) over 450 million years. The other two lineages, dominating the diagram, branched often and spread horizontally—as well as climbed vertically. Their branching and horizontal spread represented the exploration of different niches by newly evolved forms. So there it all was: evolution and the origins of diversity.
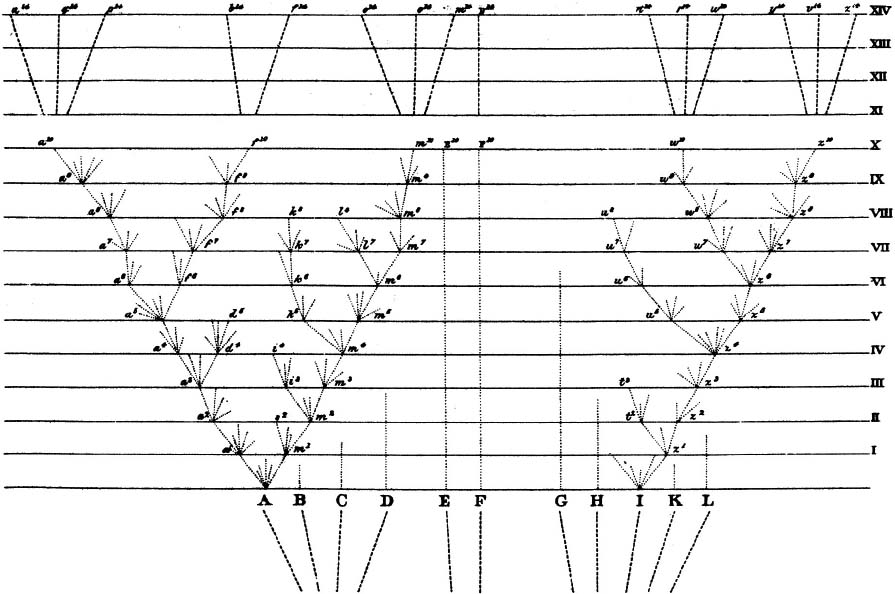
Darwin’s diagram of divergence, from On the Origin of Species, 1859.
Back in Massachusetts, Edward Hitchcock read Darwin’s book, and it stuck in his craw. This wasn’t his first exposure to the idea of transmutation (he knew of Lamarck’s work and some other wild speculations), but it was the latest statement of that idea, the most concrete and logical, and therefore the most dangerously persuasive. Like some other pious scientists who chose to see God’s hand acting directly in the fossil record—Louis Agassiz at Harvard, François Jules Pictet in Geneva, and Adam Sedgwick, who had been Darwin’s mentor in geology at Cambridge—Hitchcock wasn’t pleased.
Into the 1860 edition of his Elementary Geology, he inserted his rejoinder to Darwin’s book, based mainly on proof by authority. He noted that Pictet saw no evidence for transmutation in the fossil record of fishes. Agassiz said that the resemblances among animals derive from—where?—the mind of the Creator. “It is well to take heed to the opinions of such masters in science,” Hitchcock wrote, “when so many, with Darwin at their head, are inclined to adopt the doctrine of gradual transmutation in species.”
That was mild but firm, a dismissive shrug. Hitchcock would ignore Charles Darwin and encourage his readers to do likewise. More telling, more defensive, was his other response: he removed the trees figure from his own book. No more Paleontological Chart. It seems never to have appeared in another edition of Elementary Geology.
Darwin and Darwin’s followers owned the tree image now. It would remain the best graphic representation of life’s history, evolution through time, the origins of diversity and adaptation, until the late twentieth century. And then rather suddenly a small group of scientists would discover: oops, no, it’s wrong.
9
Molecular phylogenetics, the study of evolutionary relatedness using molecules as evidence, began with a suggestion by Francis Crick, in 1958, offered passingly in an important paper devoted to something else. That was characteristic of Crick—so brilliant and recklessly imaginative that he sometimes influenced the course of biology even with his elbows.
You know Crick’s name from the most famous triumph of his life: solving the structure of the DNA molecule, with his young American partner James Watson, in 1953, for which he and Watson and one other scientist would eventually, in 1962, receive the Nobel Prize. Crick wasn’t wasting his time, in 1958, mooning about dreams of glory in Stockholm. He was still interested in DNA, but he had moved on from the sheer structural question to other big problems. He had bent his mind intensely, but with his usual sense of merry play, to the challenge of deciphering the genetic code.
The code, as you’ve heard many times but might need reminding, is written in an alphabet of four letters, each letter representing a component—a nucleotide base, in chemistry lingo—of the DNA double helix. The four letters are: A (for adenine), C (cytosine), G (guanine), and T (thymine). DNA’s full moniker is deoxyribonucleic acid, of course, and it’s worth understanding why. The two helical strands of the double helix, twining around a central axis in parallel with each other, are composed of units called nucleotides, linked in a chain, each nucleotide containing a base (that’s the A, C, G, or T), a sugar (that’s the deoxyribose), and a phosphate group (that’s the acidic part). The sugar end of one nucleotide bonds to the phosphate end of the next, forming the two long helical strands. I just called them parallel, but to be more precise, those strands are antiparallel to each other, since the sugar-phosphate binding gives them directionality—a front end and a back end—and the front end of one strand aligns with the back end of the other. The nucleotide bases, linked crossways by hydrogen bonds, hold the strands together. The base A pairs with T, the base C pairs with G, forming a stable structure, like the steps in a spiral staircase. This is the nifty arrangement that Watson and Crick deduced.
It’s not just a stable structure, though. It’s a wondrously efficient one for storing, copying, and applying heritable data. When the two strands are peeled apart, the sequence of bases along one of the strands (the template strand) represents genetic information ready to be duplicated or used. Watson and Crick noted that capacity with exquisite coyness in their 1953 paper. The paper was lapidary, only a page long, as published in the journal Nature, and included a sketch. Near the end, having proposed their double helix structure and the matchup of bases, always A with T and C with G, they wrote: “It has not escaped our notice that the specific pairing we have postulated immediately suggests a possible copying mechanism for the genetic material.”
But copying that material, for hereditary continuity, was one thing. Translating it into living organisms was another. Translated how? By what steps does the information in DNA become physically animate?
This mystery leads first to proteins. There are four kinds of molecule essential to living processes—carbohydrates, lipids, nucleic acids, and proteins—often collectively called the molecules of life. Proteins might be the most versatile, serving a wide range of structural, catalyzing, and transporting functions. Their piecemeal production, and the controls on the process of building and using them, are encoded in DNA. Every protein consists of a linear chain of amino acids, folded upon itself into an elaborate secondary structure. Although about five hundred amino acids are known to chemistry, only twenty of those serve as the fundamental components of life, from which virtually all proteins are assembled. But what sequences of the four bases determine which amino acids shall be added to a chain? What combination of letters specifies leucine? What combination produces cysteine? What arrangement of A, C, G, and T delivers its meaning as glutamine? What spells tyrosine? This fundamental matter—how do bases designate aminos?—became known as “the coding problem,” to which Francis Crick addressed himself in the late 1950s. Solving it was a crucial step toward understanding how organisms grow, live, and replicate.
There were questions within questions. Do the bases work in combinations? If so, how many? Two-base clusters, selected variously from the group of four and in specified order (CT, CG, AA, and so on) would allow only sixteen combinations, not enough to code twenty amino acids. Then maybe clusters of three or more? If three (such as CTC, CGA, AAA), do those triplets overlap one another, or do they function separately, like three-letter words divided by commas? If there are commas, are there periods too? Four letters, in every possible combination of three, yield sixty-four variants. Are all sixty-four possible triplets used? If so, that implies some redundancy; different triplets coding for the same amino acid. Does the code include a way of saying “Stop”? If not, where does one gene end and another begin? Crick and others were keen to know.
Crick himself had also started thinking beyond that problem, to the question of how proteins are physically assembled from the coded information, with one amino acid brought into line after another. How does the template strand find or attract its amino acids? How do those units become linked? He wanted to learn not just the language of life—its letters, words, grammar—but also the mechanics of how it gets spoken: its equivalent of lungs, larynx, lips, and tongue.
Crick was back in England by the mid-1950s, after a sojourn in the United States, and based again at the Cavendish Laboratory in Cambridge, where he had worked with Jim Watson. He had a contract with the Medical Research Council (MRC), a government agency with some mandate for fundamental as well as medical research. Solving the DNA structure, though it had brought scientific fame to Crick and Watson and would eventually bring the Nobel Prize, provided no immediate cure for Crick’s dicey financial situation, all the more acute since the birth of his and his wife Odile’s third child. He had to work for pay: a modest salary from the MRC and whatever small change the occasional radio broadcast or popular article might bring. Now he was sharing his office, his pub lunches, his fevered conversations, and his blackboard with another scientist, Sydney Brenner, rather than with Watson. One colleague at the Cavendish, upon early acquaintance with Crick, concluded that “his method of working was to talk loudly all the time.” When not talking, or listening to Brenner, he spent his time reading scientific papers, rethinking the results of other researchers, combing through such bodies of knowledge for clues to the mysteries that engaged him. He was not an experimentalist, generating data. He was a theoretician—probably the century’s best and most intuitive in the biological sciences.
Sometime in 1957 Crick gathered his thoughts and his informed guesses on this problem—about how DNA gets translated into proteins—and in September he addressed the annual symposium of the Society for Experimental Biology, convened that year at University College London. His talk “commanded the meeting,” according to one historian, and “permanently altered the logic of biology.” The published version appeared a year later, in the society’s journal, under the simple title “On Protein Synthesis.” Another historian, Matt Ridley, in his short biography of Crick, called it “probably his most remarkable paper,” comparable to Isaac Newton’s Principia and Ludwig Wittgenstein’s Tractatus. It was a commanding presentation of insights and speculations about how proteins are built from DNA instructions. It noted the important but still-fuzzy hypothesis that RNA (ribonucleic acid), the other nucleic acid, which seemed to exist in DNA’s shadow, is somehow involved. Might RNA play a role in manufacturing proteins, possibly by helping express the order (coded by DNA) in which amino acids are linked one to another? Amid such ruminations, Crick threw off another idea, almost parenthetically: ah, by the way, these long molecules could also provide evidence for evolutionary trees.
As published in the paper: “Biologists should realize that before long we shall have a subject which might be called ‘protein taxonomy’—the study of the amino acid sequences of the proteins of an organism and the comparison of them between species.”
He didn’t use the words “molecular phylogenetics,” but that’s what he was getting at: deducing evolutionary histories from the evidence of long molecules. Comparing slightly different versions of essentially the same protein (such as hemoglobin, which transports oxygen through the blood of vertebrates), as found in one creature and another, could allow you to draw inferences about degrees of relatedness between them. Those inferences would be based on assuming that the variant hemoglobins had evolved from a common ancestral molecule and that, over time, in divergent lineages, small differences in the amino sequences would have crept in, by accident if not by selective advantage. The degree of such differences between one hemoglobin and another should correlate with the amount of time elapsed since those lineages diverged. From such data, Crick suggested, you might draw phylogenetic trees. Humans have one variant of hemoglobin, horses have another. How different? How long since we shared an ancestor with horses? It could be argued, Crick added, that protein sequences also represent the most precise observable register of the physical identity of an organism, and that “vast amounts of evolutionary information may be hidden away within them.”
Having tossed off this fertile suggestion, Crick returned in the rest of the paper to his real subject: how proteins are manufactured in cells. That was his way. A passing thought, with the heft of a beer truck. Essentially he had said: Look, I’m not pursuing this protein taxonomy business, but somebody should.
10
Somebody did, though not immediately. Seven years passed, during which several other scientists began noodling along various routes that would lead to a similar idea. Two of them were Linus Pauling and Emile Zuckerkandl, who gave their own fancy name to the enterprise—they called it “chemical paleogenetics”—and they converged on it by very different trajectories.
Zuckerkandl was a young Viennese biologist whose family had escaped Nazi Europe via Paris and Algiers. He got to America, did a master’s degree at the University of Illinois (long before Carl Woese would arrive there), then returned to Paris after the war for a doctorate. He found work at a marine laboratory on the west coast of France and studied the molting cycles of crabs, which involve a molecule analogous to hemoglobin. His interest drifted from crustacean physiology to questions at the molecular level, and he hankered to return to America. In 1957 Zuckerkandl finagled a chance to meet Pauling, who by then was a celebrated chemist with the first of his two Nobel Prizes already won. The prize had given Pauling some latitude to expand his own range of concerns, from lab chemistry at the California Institute of Technology to the wider world, and some leverage in pursuing those concerns. He had two in particular: genetic diseases such as sickle cell anemia and the threats posed by thermonuclear weapons, including radioactive fallout from testing. By the late 1950s, Pauling was raising his voice. He initiated a petition against atmospheric nuclear testing that more than eleven thousand scientists signed. He had become, along with Bertrand Russell, the provocative British philosopher, also a Nobel winner, one of the world’s most august peaceniks.
Pauling’s initial encounter with Zuckerkandl coincided with his increasing interest in genetics, evolution, and mutation—most pointedly, the mutations that might be caused by radiation released in weapons tests. His interest in disease led in the same direction, because sickle cell anemia is a problem that results from mutations in one of the genes for hemoglobin. Pauling found Zuckerkandl impressive enough that he offered the younger man a postdoctoral fellowship in chemistry at Caltech. Then, when Zuckerkandl arrived in Pasadena, intending to continue work on the crab-molting molecule, Pauling discouraged that project and said, “Why don’t you work on hemoglobin?”
Pauling suggested further that he take up a newly invented technique—still primitive but promising—that employed electrophoresis (separating molecules by their sizes, using electrical charge) and other methods to “fingerprint” such proteins, distinguishing one variant from another. Comparing protein molecules that way, Pauling figured, might allow researchers to draw some evolutionary conclusions. So Zuckerkandl went to work, learning the technique and applying it to hemoglobin in variant forms. Before long, he could see the close similarity between human hemoglobin and chimpanzee hemoglobin, and that human hemoglobin was less similar to hemoglobin found in orangutans. He could also tell a pig from a shark just by looking at the molecular fingerprints. Of course, there were easier ways to tell a pig from a shark, but never mind. Although it wasn’t such a precise methodology as he might have wished, this sort of molecular comparison was a start.
Over the next half dozen years, Zuckerkandl’s work thrived, and he published a series of papers with Pauling. Some of those were invited contributions to celebratory volumes, Festschriften, in honor of eminent scientists, generally on some occasion such as retirement or a big, round birthday. Such invitations came often because of Pauling’s own eminence, and he recruited Zuckerkandl as coauthor to do much of the thinking and most of the writing. In the meantime, Pauling won his second Nobel, this time the Peace Prize in recognition of his efforts against nuclear weapons proliferation and testing. That one didn’t add to his scientific reputation (in fact, he resigned from his Caltech professorship because university administrators and trustees disapproved of his peace activism), but it certainly helped amplify his public voice. He was a busy man, much in demand. The invitations—to speak, to visit, to contribute scientific papers for ceremonial volumes—continued. Because such papers didn’t normally go through the peer-review filter, they could be a little more bold and speculative than a typical journal article. One of them, written in 1963 to honor a Russian scientist on his seventieth birthday, was titled “Molecules as Documents of Evolutionary History.” Two years later, it was reprinted in English in the Journal of Theoretical Biology, giving it much broader reach and influence. Pauling and Zuckerkandl were wading into the same pond where Francis Crick had dipped his toe.