
Половой отбор: теория или миф? Полевая зоология против кабинетного знания
В пользу такого заключения говорит, в частности, степень цитируемости книги Андерссона: примерно 11 300 000 ссылок в Интернете менее чем за полсекунды. Создается впечатление, что ссылка на эту работу в начале каждой статьи служит для нее паролем принадлежности автора к большей части научного сообщества, которая движется в «правильном направлении современного мейнстрима». Опасность подобной канализированности взглядов состоит в том, что теряется ощущение возможности каких-либо иных объяснений происходящего.
Иные варианты объяснений эволюции полового диморфизма. К счастью, любознательность ученых во все времена противостояла конформизму нормальной науки, оставаясь главным инструментом в продвижении по пути непредвзятого знания. В 1970-х гг. в среде кабинетных теоретиков родилась идея конфликта репродуктивных интересов между особями противоположного пола (см., напр., Parker, 1979). Позже на этой почве сформулировали гипотезу «антагонистической коэволюции полов». Суть ее в том, что такой конфликт интересов может вести к циклическому процессу типа соревнования вооружений, как это происходит в развитии военной техники: усиление мощи брони влечет за собой возрастание разрушительной силы снарядов. Если говорить о таких морфологических структурах самцов и самок, как их половые аппараты, то такая коэволюция должна, теоретически, приводить к соответствию тех и других по принципу ключа и замка. Считают, что подобные скоррелированные преобразования признаков самцов и самок происходят сравнительно быстро в эволюционном масштабе времени[15].
В чем же отличия этой гипотезы от традиционных представлений, согласно которым половой отбор обязан выбору самками качественных самцов в ущерб некачественным? Посмотрим, что пишет по этому поводу В. Эберхард, работы которого лежат, все же, в русле парадигмы полового отбора. «Традиционный взгляд состоит в том, что издержки (costs) самки в момент ее выбора, если таковые имеют место, в среднем компенсируются для нее выгодами (benefits) от увеличения числа и качества ее потомков». Что касается идей, основанных на представлениях о конфликте полов, то здесь «… напротив, самка противостоит некоторым самцам, чтобы избежать издержек (costs), которые могли бы быть следствием нежелательного ухаживания за ней кого-либо из них и копуляции с ним. [Эти издержки могут быть связаны с] усилением пресса хищников*, уменьшение продукции яиц*, кормовыми условиями*, степенью выживаемости потомства**, влиянием эктопаразитов и болезней*, генетическими дефектами отпрысков*, повреждением самцом репродуктивных органов самки** или другими влияниями со стороны самца, которые снижают репродуктивный успех самки и имеют отношение к утрате ею контроля над процессом оплодотворения. Избегание самками этих издержек ведет, таким образом, к половому отбору среди самцов*[16]» (Eberhard, 2004а: 1948).
Эберхард рассматривает эти две предельно расплывчатые словесные конструкции в качестве гипотез, предсказания которых он намерен сравнить в критическом плане на основе эмпирических данных. В качестве объекта рассматриваются, с одной стороны, морфология органов, которые у самцов многих видов беспозвоночных служат для захвата самки в преддверии копуляции и ее осеменения, а с другой – те участки тела самки, с которыми эти органы самцов входят в контакт при таком взаимодействии партнеров. В процессе исследования были проанализированы виды, относящиеся к 61 семейству, преимущественно насекомых и пауков, а также некоторые виды клещей и ракообразных. Полученные результаты полностью опровергают предположение, согласно которому антагонистическая коэволюция приводит к развитию у самок структур, способных защитить их носителей от насилия со стороны самцов. Такие структуры не обнаружены ни у одного из 106 видов, относящихся к 84 таксонам. Более того, в 53.8 % случаев те участки тела самок, с которыми контактируют исследованные органы самцов, не показали ни малейших признаков каких-либо морфологических трансформаций.
В этой работе поражает контраст между той ее частью, которую можно было бы (условно) назвать теоретической, и другой, содержащей материалы эмпирического исследования. Если говорить о первой, то едва ли можно было найти лучший пример расплывчатости понятий, облеченных в рыхлую шелуху многословия, что является наиболее характерной чертой построений современной «теории полового отбора». Что же касается эмпирической части, то она может служить превосходным образцом сравнительного подхода в функциональной морфологии.
Как было сказано, Эберхард намеревался на этом примере сопоставить весомость двух точек зрения, именуемых им «гипотезами». Но поскольку ни одна из них не дает четких предсказаний относительно того, лежит ли в основе наблюдаемых событий половой отбор (и потому не может рассматриваться в качестве полноценной гипотезы), поставленная этим автором задача невыполнима в принципе. Как писал Р. Левонтин (1978: 25), «Если переменные состояний или параметры, на которых построена теория, измерить невозможно или если измерение сопряжено с такими ошибками, что невозможно сделать выбор между альтернативными гипотезами, теория становится пустым упражнением в формальной логике, никак не связанным с реальным миром. Теория не объясняет ничего, потому что она объясняет все» (курсив мой – Е.П.).
Интересно посмотреть, как сам Эберхард интерпретирует полученные им результаты. Он пишет: «Отбор, проистекающий из выбора самок, не исключает селекцию на основе полового антагонизма, и наоборот. Оба фактора, теоретически, могут воздействовать на одну и ту же структуру в одно и то же время, они могут взаимодействовать и улучшать эффекты действия каждого из них. Они могут также действовать последовательно, так что влияние антагонистического отбора дает толчок для вступления в силу фишеровского выбора самцов самками. Установить детальный ход истории действия двух типов отбора, исходя из ныне существующих морфологических структур, не представляется возможным. Я утверждаю, что многие данные [проведенного морфологического анализа] не отвечают предсказаниям гипотезы антагонистической коэволюции. Это говорит о том, что этот фактор не может сам по себе объяснить быструю дивергентную эволюцию рассмотренных генитальных структур самцов[17] и тех, что используются ими для захвата самок. Это не означает, однако, что антагонистический отбор не работает никогда. Гипотеза, тестируемая мной, относительно проста и рассматривается в ее крайней форме. То, что ее не удалось подтвердить, не отрицает возможности существования более сложных и, возможно, более реалистических версий гипотезы» (Eberhard, 2004а: 1956; курсив мой – Е.П.).
Итак, мы видим, что даже при попытке тестировать идею полового отбора на органах, допускающих строгое структурное описание, исторические причины их формирования не поддаются рациональному объяснению в терминах этих представлений. А что тогда можно сказать в этом плане, например, о формах брачного поведения, состоящих из множества двигательных реакций и организованных во времени таким образом, что они с трудом поддаются строгому описанию (как например, брачные демонстрации самца павлина – см. Панов, 2012а и ниже,)? Для меня совершенно очевидно, что «теория полового отбора» здесь совершенно бессильна, что я попытаюсь подробно аргументировать в разделе 9.3 главы 9.
Всю сложность проблемы, если взглянуть на нее с этой стороны, замечает и сам Эберхард. Он пишет: «Когда выбор самки осуществляется на основе стимуляции со стороны самца, критерии, используемые ей при этом, могут быть не столь легко выявляемыми (cryptic). Сюда относятся характер распределения на теле самки ее сенсорных органов, а также количество и тип синапсов между нейронами, отвечающими за реакции на действия самца, и прочие особенности ее нервной системы» (Eberhard, 2004а: 1949; курсив мой – Е.П.).
Состояние дел сегодня. Четверть века назад Андерссон, цитируя Фишера, писал: «“Из всех направлений биологии, которые наметили для нас бессмертные труды Дарвина, лишь очень немногие (few, if any), оказались столь привлекательными в качестве объекта исследований, как половой отбор”. Эту позицию Фишера (Fisher, 1915), высказанную в одной из наиболее оригинальных работ о половом отборе после Дарвина (Darwin, 1871), в наши дни, несомненно, разделяют многие биологи» (Andersson, 1994: xv). Сегодня в этой цитате слово «многие» можно смело заменить другими: «почти все, за очень редкими исключениями» (см. Заключение).
Время от времени раздаются лишь робкие возражения против самой идеи, приобретшей за последние тридцать лет характер повальной моды. О каких бы различиях между полами ни шла речь, их склонны приписывать половому отбору. Среди противников этой доктрины только единицы критикуют ее, делая упор на шаткость исходных оснований. Среди них назову Рут Хабберд, которая подчеркивает антропоморфические и андроцентрические истоки взглядов Дарвина на этот предмет (см. выше). Немногие другие возражения едва ли можно назвать убедительными, поскольку они борются скорее с симптомами, нежели с причиной болезни. Таково предложение отдельных групп исследователей заменить идеологию полового отбора другой, основанной на теории игр (Roughgarden et al., 2006) или же усилить ее, взяв другой вариант последней (Dali et al., 2006; McNamara et al., 2006). В обоих случаях это значило бы уйти от одного способа формального математического моделирования эволюции полового диморфизма, в широком смысле этого понятия («фишеровский половой отбор») к другому, примерно такого же умозрительного свойства.
Для полноты картины следует упомянуть позицию некоторых исследователей, которые акцентируют приоритет защитной, в частности, антихищнической, функции экстравагантных паттернов во внешней морфологии животных (формы, окраски и пр.) в ущерб ее значимости в межполовых отношениях (напр., Ruxton et al., 2004). Едва ли такое противопоставление правомерно, поскольку обе функции могут быть одинаково важны.
Точка зрения автора этой книги. Кратко сформулирую свое отношение к господствующим ныне взглядам на те явления, которые принято относить к сфере «полового отбора». В основе интересующих нас представлений лежит достаточно простая идея, которая, по сути дела, ничем не отличается от основополагающих в концепции естественного отбора: носители полезных признаков в силу своего повышенного успеха в воспроизведении потомства будут передавать эти признаки по череде поколений, а сами такие признаки будут все лучше выполнять свою адаптивную функция. Иными словами, ход эволюции, адаптивной по определению в рамках такого подхода, неизменно следует закону больших чисел, и с этой точки зрения может быть отображен в математических моделях.
В качестве неоспоримого довода в пользу существование полового отбора для зоологов-современников, как и для Дарвина, служат реальные факты гипертрофированного полового диморфизма, а также хорошо обоснованное эмпирически явление дифференциального репродуктивного успеха самцов (главы 4 и 8, соответственно). Однако в качестве единственно возможного объяснения эволюционного становления и того, и другого стало хорошим тоном считать половой отбор. При этом не только необходимым, но и достаточным доказательством именно этого хода событий считают выводы абстрактного характера, которые вытекают из упрощенческих, по сути дела, математических моделей. Недаром вся эта доктрина сегодня зачастую именуется не «дарвиновским», а «фишеровским половым отбором». Помимо модели Фишера, разработано множество других моделей того же характера (см. Заключение, 2). Это ведет, в частности, к тому, что обсуждение базисных вопросов теории постепенно уходит в тишь кабинетов, где идет острая конкуренция между разными формальными схемами (см. напр., Dominey, 1983). Теория начинает жить собственной жизнью.
Зоолог-практик, занимаясь исследованием своего объекта, не в состоянии не только критически оценить суть такого рода разногласий между математиками, но даже просто прочитать описанное ими на языке алгебраических формул. Поэтому он, волей-неволей, вынужден принимать «доказательства» справедливости теории на веру.
Это одна из причин единомыслия относительно ее исходных посылок. Оно естественным образом поддерживается внутри парадигмы, которой ученый бессознательно подчиняется и в рамках которой работает (Kuhn, 1962). Занимаясь своими исследованиями в сфере «нормальной науки», ученый и не нуждается субъективно в подобной рефлексии. Другие члены сообщества также не ожидают этого от коллег, поскольку разделяют те же взгляды. Кун специально подчеркивает этот факт единства мнений, которое может приводить к быстрому прогрессу «нормальной науки». В ней то, что можно условно назвать «инновациями», рождается в исследованиях, проводимых в соответствии с принятыми правилами и выработанными стандартами. Это новое знание, однако, остается преемственным по отношению к установленным теоретическим конструкциям и удовлетворяет ожиданиям, которые соответствуют формулировкам, принятым в данном научном сообществе в данное время. При этом истинная креативность, как источник попыток уйти от привычных стереотипов мышления, оказывается антагонистичной устоявшемуся коллективному сознанию (Bennett, 1968: 237). Таковы, на мой взгляд, социально-психологические и гносеологические истоки современной моды на половой отбор.
В следующей главе я обрисую формально логические и эмпирические обоснования современной «теории полового отбора», наметив истинное место изучаемых ею явлений в более широкой перспективе выяснения общих принципов сопряженной эволюции разных биологических систем (таких, например, как специализация паразитоидных насекомых к конкретным видам хозяев). На естественный вопрос, что я собираюсь противопоставить современным взглядам относительно полового отбора, ответ будет таков: изучение хода эволюции полового диморфизма вообще и эксцессивных структур, в частности, на основе данных по реконструкции филогении конкретных групп организмов.
Глава 1
«Теория» полового отбора
В наши дни, когда биология достигла таких успехов, как расшифровка генетического кода, секвенирование полных геномов ряда модельных видов (включая человека), возможности реконструкции хода филогенеза с использование методов кладистики и геногеографии, как-то даже неловко пересказывать наивные, умозрительные «догадки» Дарвина по поводу эволюции полового диморфизма у животных, ведущей, как он полагал, к прогрессивному развитию эксцессивных структур у самцов. На мой взгляд, эти построения должны были бы представляют сегодня лишь чисто исторический интерес. Однако, чтобы показать далее шаткость этих воззрений, я вынужден обрисовать их структуру в том виде, как она выглядит сейчас. При этом мне было достаточно отталкиваться от книги М. Андерссона, в которой он постарался придать расплывчатым идеям классика форму достоверного знания.
Этой попытке Андерссона в наибольшей степени способствовал обзор существующих генетико-математических моделей, которые служат, как он полагает, достаточно весомым доводом в пользу реальности полового отбора (главы 2 и 3 его книги «Половой отбор»). И если первым таким «доказательством» существования и значимости этого процесса для многих послужила модель Фишера, то вторым ныне бесспорно является книга самого Андерссона, которая непременно цитируется в начале каждой выходящей в свет статьи на эту тему. Далее, опираясь на эту сводку, я рассмотрю главные постулаты современной доктрины полового отбора.
1.1. Основные постулаты «теории»
I. Отбор половой есть нечто иное, чем отбор естественный. Вот что по этому поводу писал сам Дарвин: «Если оба пола ведут совершенно одинаковый образ жизни, и все-таки самец обладает органами чувств и перемещения, выше развитыми, чему самки[18], то может случиться, что усовершенствование этих органов необходимо самцу для отыскивания самки; но в огромном большинстве случаев такие органы служат лишь для доставления преимуществ одному самцу над другим, потому что за достаточно долгий промежуток времени, менее одаренные самцы могли бы успешно спариваться с самками; а судя по строению самки, самцы эти во всех прочих отношениях одинаково хорошо приспособлены к обычному образу жизни. Так как в подобных случаях самцы прибрели свое нынешнее строение потому, что они лучше приспособлены к переживанию в борьбе за существование, но по той причине, что это преимущество было передано исключительно мужскому потомству[19], половой подбор должен был здесь играть роль. Важность этого различия и привела меня к обозначению этой формы подбора именем «половой». Далее, таким же образом, если главная польза, оказываемая самцу его хватательными органами[20], состоит в предупреждении побега самки до прибыли других самцов или до нападения с их стороны, то и эти органы должны были улучшиться путем полового подбора, т. е. благодаря преимуществу, приобретенному известными особями над своими соперниками. Но в большей части случаев этого рода, невозможно провести различия между действием естественного и полового подбора» (Дарвин, 1896: 197–198; курсив мой – Е.П.).
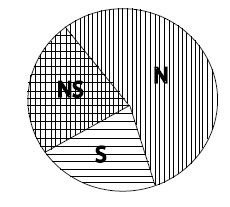
Рис. 1.1. Соотношение между естественным и половым отбором, как оно видится в доктрине полового отбора.
Из: Andersson, 1994.
Сказанное здесь, по-видимому, не кажется важным для Андерссона. Так или иначе, он ставит все точки над i в этих нечетких построениях Дарвина, облекая их в категорическую форму, которая приводится в виде графической схемы. Она показана на рис. 1.1, взятом из книги «Половой отбор». В подписи к нему сказано: «Соотношения между половым и естественным отбором. Внутри круга находятся все признаки (traits), находящиеся под действием (favored by) естественного отбора в природе. В левой половине круга содержатся те из них, которые связаны с конкуренцией из-за полового партнера (половой отбор NS и S). Некоторые признаки контролируются другим (other?) естественным отбором, как и половым отбором (NS) или только другим (?) естественным отбором (сектор N). Наконец, некоторые признаки подчиняются только половому отбору (сектор S). Этот сектор содержит набор признаков, имеющих отношение только к конкуренции между самцами. Это вторичные половые признаки, снижающие выживаемость (survival), которые Дарвин отнес к сфере действия «полового отбора». Заметьте, однако, что многие признаки контролируются как половым отбором, так и другими формами (?) естественного отбора. Относительная площадь разных секторов дана условно. Рассматриваются только те признаки, которые находятся под действием отбора в природных условиях, а не контролируются искусственным отбором или генетическим дрейфом» (Andersson, 1994: 8; курсив и знаки вопроса мои – Е.П.).
Легко видеть нечеткость и путанность предлагаемой здесь классификации форм отбора, что делает проведение границы между ними процедурой совершенно искусственной. Она не обоснована ничем, кроме чисто произвольного домысла. Это обстоятельство придает «теории» откровенный натурфилософский характер и тем самым ставит под удар все дальнейшие ее построения.
II. Межполовой и внутриполовой отбор. Внешнюю видимость того, что «половой отбор» есть некая реальная сущность, придает для непосвященных подразделение этой категории на два класса. Межполовой отбор осуществляется за счет селекции самками лучших самцов-производителей. Внутриполовой же идет, якобы, на почве конкуренции между самцами из-за самок. Он, как полагал еще Дарвин, должен быть особенно интенсивен у видов, у которых соотношение полов сильно сдвинуто в пользу самцов, что обостряет их конкуренцию из-за доступа к самкам. Второе понятие предложил Дж. Хаксли, который был склонен считать половой отбор лишь разновидностью естественного (Huxley, 1938а, б). К сожалению, от трактовок Хаксли, много более реалистичных, чем дарвиновские, в современной конструкции анализируемых взглядов сохранили одно лишь понятие внутриполового (интрасексуального) отбора (Huxley, 19386).
Сегодня теоретикам полового отбора кажется, что такое подразделение есть шаг вперед в сторону модернизации интуитивных построений Дарвина. Постулируя внутреннюю неоднородность полового отбора, они, как им кажется, уходят от чисто умозрительного (натурфилософского) взгляда на этот процесс, придавая ему видимость большей материальности и правдоподобия. В самом деле, если в явлении удается выявить хотя бы две разные составляющие, появляется возможность (в данном случае скорее иллюзорная) рассматривать его аналитически, не ограничиваясь одной лишь голой констатацией реальности этого явления, как это делал Дарвин.
III. Признаки самцов, способствующие их успеху у самок, снижают приспособленность их носителей. Идея, согласно которой самцы «обычно» окрашены ярче самок и несут вторичные половые признаки в виде всевозможных «украшений» и прочих эксцессивных структур (таких, например, как рога у многих копытных), стала главной в формулировании принципа полового отбора Дарвином. Проблема, с его точки зрения, состоит в том, что половой отбор, таким образом, противодействует естественному, который всегда должен работать в сторону повышения приспособления особей. В наши дни убеждение, согласно которому вторичные половые признаки самцов должны непременно снижать их выживаемость, нашла свое отражение в так называемом «принципе гандикапа» (Zahavi, 1975, 1993; Zahavi, Zahavi, 1997).
Суть этой идеи в том, что если уж самец смог выжить, несмотря на свою заметность, то его орнаментация означает для самки «честный» сигнал о его высоком качестве как производителя потомства.
Одно из отличий «честного» сигнала от всех прочих состоит, как полагают, в следующем. Все те структуры, которые рассматриваются в качестве украшений самца и при этом представлены в двойном наборе – справа и слева, как например, рога у копытных, должны, согласно теории, быть более или менее асимметричными. Это предположение базируется на том основании, что такие структуры есть результат интенсивного направленного[21] (strong directional) полового отбора. А тот должен, по мнению теоретиков этого направления, работать против генетических модификаторов, канализирующих развитие организма, в силу чего нарушается его стабильность. Именно это и должно приводить к неодинаковой скорости роста структур, обязанных действию полового отбора, в отличие от других, сугубо функционального характера (ноги, крылья и пр.[22]). Явление получило название «варьирующей асимметрия» (fluctuating asymmetry).
Варьирующей (флюктуирущей) она названа потому, что степень ее выраженности не одинакова у особей данной популяции (в том числе и самцов), причем отклонения от идеальной симметрии носят случайный характер (Parsons, 1990; подобнее см. раздел 10.3). Полагают, что у индивидов «высокого генетического качества» в период их онтогенеза некая буферная система должна работать против затрат (costs), тормозящих развитие ярко выраженных (elaborate) вторичных половых признаков, чего нет у генетически низкокачественных особей. В результате, высококачественные самцы будут иметь более крупные и более симметричные «честные» сигнальные структуры, а низкокачественные – меньшие по размерам и асимметричные (см., напр., Moller, Pomiankowski, 1993: 465). Все эти построения носят название «гипотезы честной сигнализации». В действительности, как нетрудно видеть, перед нами нанизывание друг на друга как минимум трех достаточно произвольных предположений, что вообще характерно для структуры рассматриваемой «теории».
Полагают также, что ярко выраженные вторично половые признак самцов могут служить честным сигналом для самок относительно качественности самца как носителя «хороших генов», поскольку это указывает ей на резистетность носителя таких признаков к паразитам. Согласно этой точке зрения, самцы с броской внешностью не только устойчивы против паразитов, но также, по сравнению с другими самцами, более изощрены в брачных демонстрациях, и/или обладают более разнообразным репертуаром вокализации, используемой при межполовых взаимодействиях. Коль скоро резистентность к паразитам, как принято думать, детерминирована генетически, самка, остановившая выбор на таком самце, может повысить жизнеспособность своих потомков. Таким образом, яркость брачного наряда самцов есть одновременно результат полового отбора и фактор дальнейшего усиления экстравагантности вторичных половых признаков, о которых идет речь (Hamilton, Zuk, 1982).
IV. Если для украшенного самца половой отбор чреват потерями (cost) в выживаемости, то самки при спаривании с ним выигрывают в том или ином отношении. Считается, что выигрыш (benefit) самки может быть «прямым» и/или «косвенным». В первом случае она приносит большее число потомков, во втором – ее отпрыски оказываются более качественными, нежели при спаривании с самцом с менее выраженными вторично-половыми признаками.