
В поисках общей теории роста человечества
Совершенно ненужную путаницу по теме сингулярности вносит так называемая «концепция Панова». Физик ядерщик А.Д. Панов, развивая идеи антидарвиниста Г.Д. Снукса, пришел к апокалиптическим результатам, главный из которых заключается в том, что в XXI веке, кроме конца исторических циклов и завершения демографического перехода, состоится еще и кризис-кризисов с «4-х миллиардолетней историей накопления противоречий».
Этот кризис-кризисов Большой истории, по мнению Панова, является переломным и завершающим моментом всей миллиардолетней истории развития жизни на Земле. Происходит он прямо сейчас (2004 год плюс-минус 15 лет) и дословно по Панову «естественным образом обобщает сингулярность человеческой истории».
Нужно ли доказывать, что биосферный кризис, назревавший миллиарды лет и разрешающийся биосферной сингулярностью в считанные годы прямо на наших глазах – всего лишь выдумка Панова? Но можно и доказать[91].
Нелепые выводы, полученные Пановым, есть результат допущенных им грубых просчетов. И, прежде всего, это физикалистский, катастрофический подход к истории, эволюции, а также навязчивое стремление уложить все реперные точки универсальной эволюции непременно в одну прогрессию.
Неловкость ситуации заключается в том, что именно Панов вводит термин «сингулярность Дьяконова», который отождествляется в его работах с несуществующей, выдуманной им биосферной сингулярностью. И сейчас уже непонятно, что он обозначает. Остается только удивляться самонадеянности и бестактности Панова, который связал результаты своих более чем сомнительных исследований с именем выдающегося историка.
Что же тогда означает сингулярность Дьяконова? (В дальнейшем будем называть ее сингулярностью Дьяконова – Капицы, т. к. С.П. Капица первым обоснованно связал ее с сингулярностью гиперболы Фёрстера.) По-видимому, за этим понятием следует оставить то смысловое значение, которое вкладывал в него сам И.М. Дьяконов, а именно: это та точка на оси времени, за которой дальнейшее сжатие исторических периодов приводит к бессмысленным результатам.
Будем пока считать, а в дальнейшем обоснуем, что она совпадает с тем моментом времени, при достижении которого эмпирические гиперболы Фёрстера и Хорнера «уходят на бесконечность», т. е. с 2022–2027 годом. Теперь, когда внесена ясность в понимание термина «сингулярность Дьяконова – Капицы», можно сформулировать еще один вопрос загадочной демографии:
Почему два процесса, продолжавшиеся тысячелетиями и напрямую никак не связанные: гиперболический рост населения Земли и сжимающиеся по закону прогрессии циклы эволюции и истории завершаются практически одновременно, а именно: конец первого цикла демографического перехода совпадает с сингулярностью Дьяконова – Капицы?
Загадки теории Капицы, несостоятельность теории Коротаева
Важнейшим этапом в развитии теоретической демографии стала феноменологическая теория, предложенная С.П. Капицей [3,4]. От эмпирического закона роста численности населения Земли от времени был сделан переход к простому дифференциальному уравнению, описывающему зависимость скорости роста от численности:
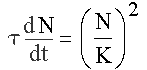
Рис. 1. Уравнение Капицы; τ = 42 года, K = (C/τ)0.5 = 67000 – безразмерная константа роста.
Из этого закона квадратичного роста скорости роста численности от численности вытекает закон гиперболического роста численности от времени. Анализируя процесс в области демографического перехода и вблизи нее, когда гиперболический рост еще не закончился, С.П. Капица немного модифицировал уравнение, описывающее рост, и ввел две константы τ и К вместо одной С (постоянной Фёрстера).
В результате удалось получить хорошую аппроксимацию зависимости численности населения мира от времени N(t) для всей истории развития человечества на протяжении более четырех миллионов лет, включая демографический переход. Были получены новые результаты, касающиеся циклов исторического развития, подсчитано количества людей, когда-либо живших на Земле. Кроме того, был предложен принцип демографического императива, согласно которому рост численности населения Земли на протяжении всей истории развития человечества не зависел ни от каких ресурсов, а определялся только самой этой численностью.
Феномен квадратичной зависимости скорости роста численности населения мира от численности С.П. Капица объясняет прямым выражением информационной природы развития, присущей только человеку. Константа К = 67000 – это численность той группы людей, которая задает характер т. н. коллективного взаимодействия Капицы. Константа τ = 42 года – «время, определяемое внутренней предельной способностью системы человечества и человека к развитию», согласно определению, которое дал этому масштабу времени С.П. Капица.
Но согласно существующим представлениям человечество долгое время не представляло собой системы, и рост населения Европы, Америки, Азии… происходил в основном независимо. Как это можно объяснить, исходя из теории Капицы?
«Вплоть до самого недавнего времени (а в особенности до 1492 г.) человечество не представляло собой системы ни в каком реальном смысле, ибо, например, рост населения таких регионов, как Старый Свет, Новый Свет, Австралия и Тасмания или Гавайские острова происходил практически полностью независимо друг от друга. Так, представляется вполне очевидным, что бурные демографические процессы, происходившие в I тыс. н. э. в Евразии, не оказали абсолютно никакого влияния на синхронную демографическую динамику, скажем, обитателей Тасмании (да и обратное влияние также было просто нулевым).
Давно ли по историческим меркам европейцы и азиаты узнали о жителях Западного полушария? Как такое человечество могло быть единым информационным полем? Вряд ли Капица допускает, что песни бардов и рассказы стариков у семейного очага при отсутствии спутниковых ретрансляторов звучали на всю Ойкумену. А если бы и звучали, то на языке, непонятном для подавляющей части Ойкумены» Ю.В. Шишков, «Демографические похождения физика».
Какова природа введенных постоянных? Какой масштаб времени задает постоянная τ? Она характеризует человека, но человек за два миллиона лет эволюции прошел путь от полуобезьяны до субтильного интеллигента, а постоянная, описывающая рост численности, не изменилась.
Почему длительность демографического перехода равна 2τ? – Нет ответа. А безразмерная константа К, столь близкая к круглому числу 216, в чем ее смысл? Размер группы, в которой проявляются коллективные признаки сообщества людей? Но для какого момента исторического времени? – Вопрос без ответа.
Само же уравнение предполагает пропорциональность скорости роста населения Земли числу связей между группами людей численностью 67 тыс. каждая. Однако, если каждая из постоянных К и τ имеет фундаментальный смысл, а не является величиной, полученной в результате статистического усреднения, то это уравнение есть не что иное, как констатация равенства приращения численности за время τ – числу связей между группами людей, численностью 67 тыс. каждая, умноженному на два.
И это равенство справедливо на огромном промежутке времени, не зависит ни от уровня развития самого человека, ни от уровня развития его технологий, ни от множества прочих факторов. Это самое большое чудо из всех рассмотренных.
Теория Капицы привлекла внимание ряда отечественных ученых, но все попытки ее развить, представить собственное видение проблемы отмечены печатью деградации. Так, А.В. Подлазов считает введение постоянных К и τ ошибочным:
«При этом уравнение (4), на наш взгляд, может быть про интерпретировано единственным образом: рост численности человечества определяется парным взаимодействием городов! Явственно присутствующий в этом положении элемент мистики обусловлен совершенно искусственным выделением одного из уровней иерархии организации общества.
Людские объединения в высшей степени масштабируемы и способны к самодостаточному существованию при численности как в десятки, так и миллионы человек. Таким образом, расщепление величины C на две константы K и τ является ошибочным и вызвано отсутствием понимания физического смысла выражения, стоящего в правой части уравнения…» [5].
А. Коротаев, Н. Комарова, Д. Халтурина [7], реставрируя модель М. Кремера с «мальтузианско-кузнецианским» уклоном, подгоняют свою «компактную» систему дифференциальных уравнений к желаемому результату. А именно: скорость роста численности населения мира пропорциональна квадрату этой численности. Константы Капицы К и τ в эту систему не входят, возвращена постоянная Фёрстера. Вся логика построена на рассуждениях вида А ~ В, А ~ С, следовательно, А ~ В·С. Исходные линейные зависимости считаются очевидными:
«Модель М. Кремера дает этому очень убедительное объяснение (хотя сам М. Кремер и не показал этого в достаточно ясном виде). А объяснение это заключается в том, что рост численности населения мира с 10 до 100 млн человек подразумевает, что и уровень развития жизнеобеспечивающих технологий вырос приблизительно в десять раз (так как он оказывается в состоянии поддержать существование на порядок большего числа людей). С другой стороны, десятикратный рост численности населения означает и десятикратный рост числа потенциальных изобретателей, а значит, и десятикратное возрастание относительных темпов технологического роста.
Таким образом, абсолютная скорость технологического роста вырастет в 10*10 = 100 раз… А так как N стремится к технологически обусловленному потолку несущей способности Земли, мы имеем все основания предполагать, что и абсолютная скорость роста населения мира (dN/dt) в таком случае в тенденции вырастет в 100 раз, то есть будет расти пропорционально квадрату численности населения» [7].
Изобретательская теория Коротаева и соавторов требует большого числа незначительных изобретений. На самом же деле новационный (и инновационный) процесс устроен иначе: все действительно значимые изобретения, открытия немногочисленны и представляют собой цепочку, в которой каждое последующее звено вытекает из предыдущего. Для Мир-системы в XIX и XX веках – это так называемые «базисные инновации» (по Л. Нефедову), которые в течение последующих десятилетий играют роль локомотива мировой экономики.
Например, в начале прошлого века Планк открывает, что процессы излучения и поглощения электромагнитной энергии нагретым телом происходят дискретно, а Эйнштейн вводит понятие кванта излучения. В двадцатых годах создается квантовая теория; в тридцатых – физика твердого тела; в конце сороковых изобретен первый транзистор; в начале шестидесятых – первая интегральная микросхема.
В конце семидесятых – первый твердотельный компьютер; в начале XXI столетия сотовый телефон становится средством массовой коммуникации. Вряд ли кто-нибудь будет возражать, что изобретение сотовой связи очень сильно повлияло на социум, в том числе и в плане роста его численности.
Но Макс Планк сделал свое открытие в известной мере случайно, оно могло быть совершено другим исследователем как раньше, так и позже отмеченного момента времени. И если сдвигать это первое звено во времени, то с ним сдвигается и вся цепочка. Именно так, а не по Коротаеву, когда мелкие инновации «мгновенно» поднимают потолок несущей способности Земли, устроен научно-технический, социальный и демографический прогресс.
В статье «Человечество подошло к пределу своего роста» А.В. Коротаев и соавторы с удивлением замечают, что модель Кремера заводит их в тупик. Ведь после демографического перехода рост численности населения Земли должен полностью прекратиться, а значит прекратится и всякий творческий процесс. Творчество больше не нужно? – Вопрошают они. А, может, все-таки модель Кремера неверна? Ведь сам Кремер ее так до конца и не сформулировал. Что-то, видимо, его удержало.
Развивая «мальтузианско-кузнецианский» подход, авторы [7] формулируют задачу на языке кибернетики и вводят в рассмотрение нелинейные обратные связи между основными субсистемами «Мир-системы». Но все попытки объяснить как гиперболический рост, так и демографический переход положительными и отрицательными обратными связями в «Мир-системе» (для человечества в целом!) чисто умозрительны, разноплановы и неубедительны. Можно ли поверить в то, что «положительная обратная связь второго порядка», в случае роста численности народонаселения, столь сбалансированна и точна, что погрешность в формуле Фёрстера для показателя степенной функции составляет всего один процент? (Точнее, n = -0,99 ± 0,009).
После демографического перехода уже два контура обратной связи, положительной и отрицательной, т. е. целая система автоматического регулирования, удерживают численность на фиксированном уровне. При этом речь не идет об исчерпании каких бы то ни было ресурсов. Несущая способность Земли может выдержать значительно большую численность. И совершенно непонятно какие такие ограничения механизма развития начинают вдруг играть доминирующую роль.
Законы роста численности изолированных популяций
Введение
Популяция – это совокупность особей одного вида, обладающая общим генофондом и проживающая на общей территории. Она является элементарной генетической единицей вида, первой надорганизменной биологической системой. Считается, что любая популяция способна к неопределенно долгому самостоятельному развитию.
Биотическим потенциалом вида называется показатель скорости роста численности особей этого вида при отсутствии ограничивающих факторов. Совокупность же таких ограничивающих рост популяции факторов называется сопротивлением среды.
Состояние равновесия между биотическим потенциалом вида и сопротивлением среды, поддерживающее постоянство численности популяции, называют популяционным гомеостазом. При его нарушении возникают колебания численности. Различают периодические и непериодические колебания численности популяции.
Обычное, нормальное состояние популяции – это гомеостаз с неизменной численностью, который поддерживается отрицательными обратными связями, обеспечивающими такой гомеостаз. Но в редких случаях численность популяции меняется и за короткий промежуток времени может значительно возрасти или уменьшиться. Этот редкий случай нарушения гомеостаза только и будет здесь нас интересовать.
Причем нами будут рассмотрены только законы роста: законы, по которым растет численность изолированной популяции, т. е. популяции более или менее отделенной в пространстве от других аналогичных совокупностей того же вида. Эти законы представляют для нас интерес в связи с законом роста численности населения Земли.
Идеализации
Построение математической модели какого-либо объекта, явления неизбежно требует принятия некоторых упрощений, идеализаций. Чем больше идеализаций, тем проще модель, тем удобней с ней работать и тем уже спектр явлений, который она способна описать.
С другой стороны, идя по пути усложнения модели, нужно иметь в виду, что даже максимально сложная, «все учитывающая» модель все равно остается всего лишь моделью и неспособна полностью описать явление, зато способна перенести львиную долю внимания исследователя с самого явления на абстрактный математический аппарат, его описывающий.
Поэтому в математическом моделировании существует золотая середина степени усложнения. В математической экологии эффективны простые модели с большим количеством идеализаций. Рассмотрим идеализации для модели роста изолированной популяции, т. е. такой популяции, взаимодействия в которой возможны только между представителями данной популяции [12]:
1. Постоянство внешних условий, т. к. прежде чем исследовать роль внешних воздействий следует проанализировать свойства идеальной, изолированной популяции, на динамику численности которой влияют лишь биотические факторы, причем только те из них, что связаны с внутривидовой деятельностью. Под постоянством внешних по отношению к растущей популяции условий будем понимать также и независимость роста (при прочих равных условиях) от того, на каком участке шкалы физического времени он наблюдается. В уравнениях такого роста не должно, следовательно, явным образом присутствовать время, т. е. они должны быть автономными.
2. Целочисленное число особей популяции заменяется для удобства на непрерывную, действительную величину.
3. Рассматриваемая популяция считается однородной, т. е. полностью пренебрегается ее половой, возрастной, генотипической и какой-либо другой структурированностью[92].
4. Рост численности любой популяции есть, строго говоря, случайный процесс, который должен описываться на языке теории вероятностей. Но при исследовании изменения численности популяций с большим числом членов естественно описывать эти изменения на языке средних величин.
5. В случае неперекрывающихся поколений в дискретных моделях принимается синхронное размножение у всех организмов при достижении определенного возраста. Что хотя и не соответствует действительности, позволяет упростить математический аппарат, причем без отрицательного влияния на результат. Момент появления новой особи в непрерывных моделях считается равномерно распределенным на отрезке времени, равном среднему времени жизни особи.
6. В случае перекрывающихся поколений скорость изменения численности может определяться численностью не в текущий, а в некоторый предшествующий момент времени. Динамика изменения численности описывается здесь уравнениями с запаздывающим аргументом. Такое запаздывание, в случае если оно сравнимо или превосходит характерное время системы, может приводить к колебаниям численности и даже к резонансам: колебаниям с нарастающей амплитудой. Пренебрежение таким запаздыванием – еще одна часто принимаемая идеализация.
7. Исследуемая система предполагается либо локальной, т. е. имеющей достаточно малые размеры (для таких систем понятия численности популяции и ее плотности являются синонимами), либо постулируется полное перемешивание, когда особь за время жизни успевает побывать на всей территории обитания популяции. Для человеческого общества предполагается его информационная связность на всем протяжении роста. При исследовании локальных или сосредоточенных сообществ изучается исключительно временна́я динамика. На самом деле сосредоточенных сообществ не существует, а реальная протяженность ареала обитания популяции может в сотни и тысячи раз превышать величину перемещения особи за поколение. Модели пространственно-распределенных сообществ включают анализ как временно́й, так и пространственной организации этих сообществ. Они описываются уравнениями типа диффузия-кинетика, решение которых зачастую сопряжено с непреодолимыми математическими трудностями.
8. Рост численности изолированной популяции предполагается свободным, никем и никак не управляемым ростом, происходящим в естественных природных условиях.
* * *Условно все идеализированные модели биологических систем можно разделить на три типа: регрессионные, качественные и имитационные [11].
А. Регрессионные зависимости – это не более, чем формулы, описывающие связь различных характеристик системы, которые при этом не претендуют на какой-либо каузальный, физический или биологический смысл. Для построения регрессионной модели достаточно статистически достоверных наблюденных корреляций между переменными или параметрами системы.
Б. Качественные (базовые) модели. В любой науке существуют простые модели, которые поддаются аналитическому исследованию и обладают свойствами, позволяющими описывать целый спектр природных явлений. Их задача качественно описать систему, в данном случае растущую изолированную популяцию. Базовые модели обычно представляют собой системы дифференциальных или разностных уравнений относительно небольшой размерности, допускающие аналитическое и качественное компьютерное исследование. Эти модели позволяют ответить на вопросы: возможны ли в системе колебания, переключения режимов функционирования, пространственно-неоднородные решения, квазистохастическое поведение. При этом важно понимать, что истинные причины наблюдаемого поведения популяции, особенности роста ее численности могут никак такой моделью не отражаться.
В. Имитационные модели. По меткому выражению Р. Шеннона имитационное моделирование – это нечто промежуточное между искусством и наукой. Суть его заключается в исследовании сложной математической модели с помощью вычислительных экспериментов и обработки результатов этих экспериментов. Как правило, создатели такой имитационной модели пытаются максимально использовать всю имеющуюся информацию об объекте моделирования как количественную, так и качественную. При этом модель может получиться разной у разных авторов, поскольку точные формальные правила ее построения отсутствуют.
Целью нашего исследования является построение качественной (базовой) обобщенной модели роста численности изолированной популяции с учетом всех обозначенных здесь идеализаций.
Каузальный анализ законов роста
Каузальный анализ описывает явление на языке причинно-следственных связей. В его основе лежит стремление понять это явление на основе логики типа: «X вызывает Y». Факторы, которые вызывают какие-то изменения, называются независимыми переменными, в то время как переменные, изменяющиеся под действием этих факторов, называются зависимыми.
В общем случае присутствие причинно-следственных связей означает, что наличие изменений меняет вероятностные характеристики последствий. В чем задача каузального анализа роста численности популяции? Она заключается в поиске причин, по которым ее численность растет по тому или иному закону.
Самый простой в каузальном смысле рост – это экспоненциальный рост. Закон экспоненциального роста считается первым законом экологии популяций. Его можно уподобить первому закону Ньютона в механике. Когда на тело не действуют никакие другие тела – оно сохраняет состояние покоя или равномерного и прямолинейного движения. (Скорость растет, убывает, меняется по направлению только тогда, когда на тело действует сила.) Когда на растущую в условии изобилия ресурсов изолированную популяцию не оказывают воздействия никакие внутренние или внешние ограничения – она растет экспоненциально. Отклонение от экспоненты и, в частности, неизменная численность возможно лишь при наличия сопротивления (ускорения) со стороны среды обитания.
Причина экспоненциального роста без смертности (для делящихся микроорганизмов) заключена внутри черного ящика процесса репродукции элементарной ячейки популяции. И прирост численности здесь всегда будет пропорционален самой численности. Для популяций животных (многоклеточных организмов) разность между приростом численности за счет рождаемости и ее убылью по причине смертности за единицу времени – также пропорциональна самой численности.
В обоих случаях – это строгое равенство при выполнении принятых выше идеализаций, т. к. рост популяции здесь представляет суперпозицию не оказывающих взаимного влияния процессов. И если, скажем, увеличить численность в два раза, то и ее естественный прирост также должен возрасти в два раза. Поэтому экспоненциальный рост популяции, происходящий в естественных природных условиях, информационно и каузально прост и его можно считать причинно-самодостаточным, а сам закон экспоненциального роста – причинным.
Причинным в том смысле, что рост популяции здесь может быть представлен как автокаталитический, самоускоряющийся процесс, причиной которого является положительная обратная связь между численностью и естественным приростом, природа которой заключена в простом росте (по закону геометрической прогрессии на последовательности интервалов равной длительности) некоторого числа параллельных, в первом приближении не взаимодействующих элементарных продукционных процессов.
Самодостаточным в том смысле, что никаких других причин у этого роста кроме тех, что заключены внутри черного ящика процесса репродукции элементарной составляющей популяции – здесь нет. Если же учитывать влияние взаимодействий между членами популяции, то линейное уравнение экспоненциального роста необходимо трансформировать в нелинейное.
Примеры таких уравнений мы приведем ниже. При этом прирост численности на особь, элементарную ячейку размножающейся популяции, будет зависеть от ее общей численности.
* * *Возможны два каузальных подхода при описании такого нелинейного роста.
1. В первом подходе причина роста ищется исключительно в связях между членами популяции, при этом полностью пренебрегается составляющей прироста без учета взаимодействий, т. е. индивидуальной способностью к размножению элементарной составляющей популяции, которая при отсутствии взаимодействий вызывает экспоненциальный рост. Так, в моделях роста численности населения Земли полагают, что мировой естественный прирост пропорционален квадрату полной численности населения Земли при любых значениях этой численности.