
Временнáя структура биосистем и биологическое время

Марина Чернышева
Временнáя структура биосистем и биологическое время
Sankt-Petersburg State University
M. P.Chernysheva
TEMPORAL STRUCTURE of biosystems and biological TIME
St.Petersburg
Super Izdatelstvo
2016
Введение
Природа Времени – одна из глобальных проблем, к решению которых наука неоднократно возвращалась на протяжении всей истории ее существования. Эволюция представлений о Времени от античности до XX-го века глубоко проанализирована в классическом труде Дж. Уитроу «Естественная философия времени» (1964), в монографиях М. И. Элькина (1985), П. П. Гайденко (2006) и других авторов. Начиная с ХХ века философские аспекты этой проблемы неизменно связаны с естественнонаучными подходами к ее решению (Шредингер, 2002; Чижевский, 1973; Уинфри, 1986; Козырев, 1963, 1985, 1991; Пригожин, 2002; и др.). В работах выдающихся отечественных исследователей находим идеи, давшие начало целым направлениям в науке о времени. Так, И. М. Сеченов положил начало исследованиям по влиянию двигательной активности на субъективное время человека. И.П. Павлов, впервые описавший рефлекс на время, фактически заявил о способности мозга к запоминанию временных интервалов. Н. П. Пэрна (1925), сотрудник кафедры физиологии Петроградского университета, впервые описал ритмы ряда физиологических процессов человека. Д. И. Менделеев, описавший движение цветка вслед за изменением положения солнца, определенно продемонстрировал наличие околосуточного (циркадианного) ритма движений растений, гормональный механизм которого был описан позже (В. Н. Полевой, 1982). В работах А. А. Ухтомского прослеживается мысль о важности временного фактора в работе нервной системы и в, частности, в формировании доминанты (Ухтомский, 1966; Соколова, 2000). Один из гениев русского Ренессанса начала ХХ века, В. И. Вернадский, не только ввел рубрикацию специфического для разных систем времени (геологического, исторического, биологического, социального), но и обосновал представление о биологическом времени как основном и первичном, придав ему «космический статус» по причине способности биосистем к движению и размножению (Вернадский, 1989). Эту же особенность живых организмов подчеркивал Э. Шредингер (2002).
Наряду с мультидисциплинарными подходами к решению проблемы природы Времени (Аксенов, 2000; Вакуленко и др., 2008; Казарян, 2009; Коганов, 2009; Козырев, 1989; Коротаев, Киктенко, 2012; Лебедев, 2004; Левич, 2000, 2002, 2013; Хасанов, 2011; Чураков, 2012; Шихобалов, 2008, и др.), огромный объем исследований, начиная со второй половины ХХ века, посвящен природе биологического времени (Aschoff, 1960; Уинфри, 1990; Питтендрих, 1984; Алпатов, 2000; Романов, 2000; Оловников, 1973, 2009; Скулачев, 1995; Загускин, 2004, 2007, и др.). Достижения физики, химии, математики и биологии предопределили разработку разнообразных новых методов исследования, позволивших открыть белки часовых генов (clock-genes proteins), формирующие механизм околосуточных ритмов для многих функций организма. Важность активности clock-белков и clock-осциллятора для здоровья и адаптации человека к пространственно-временному континууму окружающей среды обусловили соответствующую тематическую направленность большинства работ современных отечественных и зарубежных исследователей. В отечественной биологии и медицине «штурм» клеточно-молекулярных механизмов биологического времени привел к выдающимся открытиям: созданию теломерно-редусомной теории контроля продолжительности жизни (Оловников, 1973, 2009) и представления о роли митохондрий в процессах старения (Скулачев, 1995), а также к развитию геронтологических аспектов роли гормонов эпифиза и тимуса (Анисимов, 2010; Хавинсон и др., 2011; Кветной и др., 2011). В работах зарубежных исследователей выявлены функции отдельных clock-белков, условия формирования clock-осциллятора и ритмов с разными темпоральными параметрами (см. Golombek et al., 2014), а также развиты представления о системах синхронизации clock-осцилляторов разных структурных уровней организма. Растущее понимание специфики клеточных, тканевых, органных и системных генераторов временных процессов определяют начинающийся возврат зарубежных авторов к «системному мышлению» в аспекте проблемы Времени (Blum et al., 2012; Mohawk et al., 2012). Заметим, что у отечественных исследователей системный подход в изучении этой проблемы всегда оставался в поле внимания (Черниговский, 1985; Баранникова и др., 2003; Кулаев, 2006; Январева и др., 2005; Журавлев, Сафонова, 2012, и др.). Наряду с очевидными успехами в изучении чувствительных к «ходу времени» (термин Н.А. Козырева) биологических объектов, остаются мало разработанными вопросы о временной структуре живых организмов, взаимосвязи клеточно-молекулярных и системных таймеров, сенсорах Времени и пока открыт вопрос о природе Времени. По мнению автора, обширный круг исследований биосистем, выполненных к настоящему времени в мире, позволяет предложить определенные решения по перечисленным вопросам.
Глава I
Биологическое время
«Понять “природу” времени, – значит указать его природный референт, т. е. процесс, явление, “носитель” в материальном мире, свойства которого могли бы быть отождествлены или корреспондированы со свойствами, приписываемыми феномену времени».
А.П. Левич, 2000.1.1. Феномен жизни
Вынесенное в эпиграф высказывание Александра Петровича Левича представляется совершенно справедливым в свете представлений Г. Лейбница и Н.А. Козырева об энергетической природе времени и его «активных свойствах». Действительно, по аналогии с историей открытия электрона по иммерсионному следу в камере Вильсона, биологические процессы, обладающие рядом темпоральных параметров и потому являющиеся по сути временными процессами, вполне могут быть «референтами» времени и отражать его воздействие. Для понимания «природы» времени в биосистемах важен анализ факторов, определяющих специфику живых организмов по сравнению с косными системами
Феномен жизни и отличия живого организма от косных систем, во все времена привлекали внимание философов и представителей естественных наук (Аристотель, 1937; Страхов, 2008; Вернадский, 1989; Ухтомский, 1966; Шредингер, 2002, и многие другие). Очевидно, что общность базисных законов природы не исключает особенностей их проявления в условиях специфики биосистемы, косной природной или искусственной систем. К их числу, в первую очередь, следует отнести законы термодинамики, определяющие для любой системы возможность и длительность работы, а также время существования (продолжительность жизни). Признавая справедливость законов термодинамики для всех объектов Вселенной, многие исследователи отмечают специфику проявлений второго начала термодинамики для живых организмов (Шредингер, 2002; Пригожин, 2002, и др.). Среди таковых, прежде всего, отмечается невозможность «тепловой смерти» для живых организмов вследствие стремления биосистем к стабилизации уровня энтропии (Вернадский, 1989; Пригожин, 2002; Пригожин, Стенгерс, 2000, и др.).
В основе жизнедеятельности биосистем лежат разнообразные процессы, использующие химическую, механическую, электрическую, световую и другие виды энергии. Как известно, при реализации различных функций (работы) в любой системе происходит частичное преобразование той или иной энергии в тепловую, которая может быть утрачена через теплорассеивание в окружающую среду или частично задержана, определяя уровень хаоса (энтропии) в структурах организма. Для живых организмов справедливы и другие известные определения энтропии: как меры степени неструктурированности потоков энергии и меры термодинамической возможности определенного состояния или процесса. Множественность возможных определений энтропии для биосистемы подчеркивает и разнообразие путей ее регуляции.
Известно, что с точки зрения термодинамики биосистемы относятся к неустойчивым открытым системам, отличающимся ростом потерь энергии, диссипатировавшей в тепловую в ходе реализации различных функций. Это согласуется, на первый взгляд, с низкими значениями к.п.д., вычисленными для мышечного сокращения, процессов фотосинтеза и т. п. Однако, этому противоречит весьма небольшая величина энтропии: для организма человека она составляет около 300 энтропийных единиц. В тепловом эквиваленте ее достаточно для испарения одного стакана воды (Опритов, 1998). В чем причина?
Согласно второму началу термодинамики, постулированному для замкнутых механических систем, самопроизвольное (то есть без участия внешних источников энергии) протекание процессов, идущих с понижением уровня энтропии, невозможно. На его основании сложилось убеждение (Шредингер, 2002; Климонтович, 1996; Пригожин, Стенгерс, 2000), что низкий уровень энтропии у живых организмов обусловлен их «самоорганизацией», возможной лишь благодаря поступлению извне энергии, компенсирующей потери, и информации (как негэнтропии). Однако современные знания о структуре и функциях живых организмов свидетельствуют о существовании у них ряда конкретных механизмов, направленных на снижение роста энтропии.
Одной из характерных черт живого организма, обеспечивающей его относительную независимость от внешних источников энергии, является множество собственных источников энергии. Так, энергия выделяется при изменении конформационной структуры белка, межмолекулярных взаимодействиях, пищеварении, сокращении мышц и многих иных биохимических и физиологических реакциях на уровне клеток, тканей, органов и организма в целом. Большинство структур организма животных и человека, – легкие, сердце, сосуды, печень, мышцы, кожа, пищеварительный тракт и другие, – участвуют в продуцировании энергии. При этом печень дает до 60 %, а скелетные мышцы при сокращении и расслаблении – до 15 % общего термогенеза организма. В основном эта энергия идет на образование новых химических связей в молекулах и при межмолекулярных взаимодействиях, на хемо-механические сопряжения в мышцах, процессы синтеза веществ и т. д., однако частично химическая и другие виды энергии диссипатируют в тепловую. Следовательно, образования организма одновременно играют роль генераторов энергии и диссипативных структур. Динамика этих процессов в клетках и жидкостных средах строго поэтапно регламентируется ферментами, что предотвращает значительные скачки температуры и роста обобщенной энтропии. Кроме того, в отличие от механических систем, тепловая энергия рассеивается не только во внешнюю, но и во внутреннюю среду, где она используется для поддержания температуры тела, активности ферментов, тонуса мышц, возбудимости нервной системы, определяет генез и силу эмоций. Этим объясняется необходимость для жизнедеятельности оптимального уровня обобщенной энтропии.
Морфо-функциональная организация дыхательной, пищеварительной и сердечно-сосудистой систем как совокупности протоков и помп на уровне организма обеспечивают градиент (давлений, рН, температур) и направленность, а также структурированность потоков энергии, что снижает скорость роста обобщенной энтропии. Этому способствует также активность хемо-, механо- и терморецепторов этих физиологических систем, которые, фактически, являются постоянным источником эндогенной информации (как негэнтропии). Это соответствует представлению об информации как основе структурированности биосистем (Левич, 1978). Поскольку рецепторы морфологически взаимосвязаны с сенсорными нейронами, эта периферическая информация поступает в итоге в головной мозг. Его структуры – другой источник эндогенной информации, хранимой в памяти, которая также может быть использована для снижения роста обобщенной энтропии организма. Свидетельством тому служит появление при гипоксии или остром нервном истощении (в случаях природных или социальных катаклизмов) сновидений и галлюцинаций как образов или эпизодов, извлекаемых из памяти (Пуховский, 2000, и др.).
Наличие эндогенных источников энергии и информации делает понятной другую важную особенность живого организма: саморегуляцию «меры открытости» его как термодинамической системы в процессе взаимодействия с окружающей средой (Чернышева, Ноздрачев, 2006; Chernysheva, 2005). Например, известно (Пастухов и др., 2000, и др.), что терморегуляция в условиях холода приводит к усилению теплоизоляции, снижению теплоотдачи и уменьшению чувствительности терморецепторов. Следовательно, стремление к гомеостазису как состоянию, близкому к стационарному, сопряжено в данном случае с направленностью функций организма на относительную «замкнутость» или изоляцию организма от окружающей среды в термодинамическом аспекте. Другим примером регулируемого организмом снижения уровня взаимодействия с окружающей средой может служить усиление интровертивности у женщин в состоянии беременности, у людей с патологиями внутренних органов или у здорового человека во сне.
Частичное использование диссипатированной энергии в качестве энтальпии для жизнедеятельности организма является одним из конкретных механизмов сопряжения термодинамически необратимых процессов, протекающих с возрастанием энтропии, и частично обратимых, реализуемых при относительно постоянном ее значении в условиях гомеостазиса. По мнению В. А. Опритова (2000) такое сопряжение является одним из «изобретений» живых организмов, позволяющих обойти «запрет» второго начала термодинамики на самопроизвольное снижение уровня энтропии. На примере известного феномена сопряжения процессов окисления с фосфорилированием у растений при фотосинтезе автор подчеркивает, что для такого сопряжения необходимо соблюдение двух условий: 1) сопрягаемые процессы должны различаться по уровню энергии, чтобы энергия могла перейти от одного процесса к другому; 2) процессы должны иметь общий компонент (-ы) (химический или структурный). Первое условие основано на том, что необратимые процессы, характерные для неустойчивых открытых систем, служат донором энергии для процессов обратимых, свойственных открытым системам в состоянии, близком к стационарному. Примеры подобного сопряжения многочисленны на всех структурных уровнях организма животных и человека – от молекулярного до системного. На уровне целостного организма в этом плане интересно сопоставление состояния гомеостазиса или относительного постоянства внутренней среды и реакции на воздействие стресс-фактора (Чернышева, 2003; Чернышева, Ноздрачев, 2006).
Состояние гомеостазиса можно рассматривать как условный аналог стационарного состояния, описанного для открытых неустойчивых механических систем как «норма хаотичности» (Климонтович, 1996; Линг, 2008). Оно характеризуется варьированием основных параметров жизнедеятельности (температуры, содержания сахара в крови, уровня тревожности и т. п.) в границах их оптимальных значений. Эти границы определяются геномом данного организма и, в свою очередь, обусловливают возможности адаптации организма к воздействиям окружающей среды. Адаптивность врожденных и выученных реакций позволяет сдерживать рост обобщенной энтропии организма на оптимально стабильном уровне. Так, при повторном воздействии на организм какого-либо фактора окружающей среды из памяти извлекаются наиболее адаптивные к нему реакции без дополнительных затрат энергии на их формирование. Это способствует сохранению относительно стабильного уровня гомеостазиса организма и оптимально минимальной скорости роста уровня энтропии. Следовательно, врожденную («геномную») память, характерную для живых организмов, а также память, формируемую индивидуально в процессе обучения, можно рассматривать как механизмы, обеспечивающие возможность относительно обратимых процессов (например, извлечения из памяти адаптивных реакций) и их сопряжения с относительно необратимыми (в силу роста энтропии) метаболическими, висцеральными и двигательными реакциями.
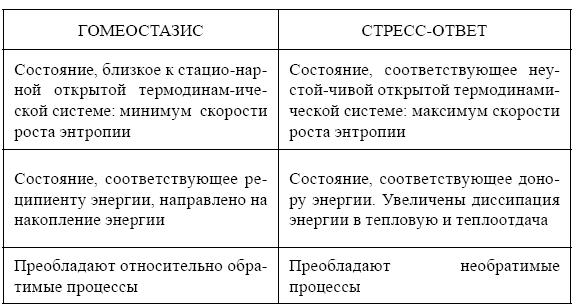
Адаптивность реакций живого организма на воздействия обусловливает целостность его структур и способствует увеличению продолжительности жизни. Воздействие экзо- или эндогенных стресс-факторов и формирование и реализация ответных адаптивных реакций, требующие затрат энергии, нарушают гомеостазис организма, повышая его термодинамическую неустойчивость. Это проявляется в характерной для стресс-реакции активации нервной, эндокринной и висцеральных систем организма, повышении уровня обмена веществ и энергии. Регистрируемые при этом повышение температуры тела, рост возбудимости нервной системы и эмоциональные реакции свидетельствуют о росте уровня обобщенной энтропии. Следовательно, термодинамический аспект стресс – ответа организма отражает его соответствие состоянию открытой системы с повышенной неустойчивостью, выполняющего функцию энергетического донора прежде всего для механизмов восстановления гомеостазиса. Это соответствует представлениям творца теории стресса Г. Селье (1960) о необходимости стресса (эустресса – стресса без патологий) для поддержания жизнедеятельности биосистем и, с другой стороны, созвучно представлению В. И. Вернадского (1989): эволюция биосферы «питается энтропией» Соотношение относительно обратимых и необратимых процессов, относительно стационарных и неустойчивых состояний при гомеостазисе и стрессе представлено в таблице I. Эти термодинамически различные процессы/состояния несомненно сопряжены не только структурно, через общие жидкостные среды и физиологические системы, но и функционально. Характерно, что в состоянии, близком к максимально неустойчивому, при стресс-ответе, активируются механизмы структурирования потоков энергии, наиболее наглядные в нервной системе.
Так, диффузная активация структур центральной нервной системы при стрессе обусловлена их связями с ретикулярной (сетчатой) формацией ствола головного мозга. Ее можно рассматривать как систему первичной структуризации энергии, выделяемой в процессе обработки информации, и, тем самым, – как механизм предупреждения дальнейшего роста энтропии. Возможно, это наиболее древняя функция нервной системы, так как в эволюции, впервые возникнув у кишечнополостных (Hydroidea), она имеет сетчатую структуру, подобно ретикулярной формации мозга позвоночных. Модельные эксперименты, проведенные на гидрах с фармакологически выключенной нервной системой, показали, что функции других систем сохраняются относительно постоянными, но укорачивается продолжительность жизни животных, что может быть следствием ускоренного роста уровня обобщенной энтропии.
В эволюции нервной системы прослеживается усиление роли и другого фактора снижения энтропии, – развития тормозных систем и увеличения разнообразия тормозных медиаторов, выделяемых нейронами и глиоцитами. На активную роль торможения в организации живого организма как биосистемы, особенно в связи с концентрацией внимания, формированием доминанты, впервые указал А. А. Ухтомский (Ухтомский, 1966).
Еще одним механизмом снижения обобщенной энтропии в живых организмах является асимметрия (Чернышева, 2003). Хотя структурно-функциональная асимметрия является одним из характерных свойств многоклеточных, но ее энергетическая «выгодность» по сравнению с симметрией была показана и на атомарном уровне (Мøller e.a., 2002): при асимметричном делении лучом лазера ядер гелия и фермия энергетический порог ниже, а суммарная кинетическая энергия осколков ядер выше, чем при симметричном.
Таблица 1.
Сравнительная термодинамическая характеристика функциональных состояний гомеостазиса и стресс-ответа организма (по: Чернышева, Ноздрачев, 2006)
В живых организмах асимметрия как феномен обладает уникальными свойствами. С одной стороны, она повышает неустойчивость организма и, следовательно, его энергетический потенциал, способствуя увеличению скорости обмена веществ и уровня энтропии, росту возбудимости и, сопряженно, – увеличению сенсорной чувствительности и объема воспринимаемой информации. Последнее, с другой стороны, определяет вклад асимметрии в сдерживание роста обобщенной энтропии. Этому способствуют и проявления морфо-функциональной асимметрии, описанные для парных структур. Например, меж-полушарная асимметрия головного мозга животных и человека (Сhernysheva, 2006; Nikolaeva, Leutin, 2011) заключается в различии не только ряда функций двух полушарий, но и уровня обмена веществ и обобщенной энтропии. Так, правое полушарие обладает более высоким уровнем обмена веществ и энергии, обобщенной энтропии, тогда как левое – более низким (Gur et al., 2002; Andrew, 2002; Чернышева, 2003). Взаимосодействие парных структур направлено на снижение энергозатрат как «цены адаптации».
Взаимосвязь асимметрии с регуляцией уровня энергообмена организма подчеркивает усиление ее при стрессе. О важности феномена свидетельствует также эволюция проявлений морфо-функциональной асимметрии на всех уровнях организации живого. Примером может служить быстрый переход в эволюции плана тела животных от пятилучевой симметрии к трехосевой асимметрии, что отражает процесс адаптации плана тела живых организмов к трехмерному окружающему пространству.
Итак, перечислим кратко свойства живых организмов, позволяющие снижать рост обобщенной энтропии вопреки второму началу термодинамики. Среди них: сопряжение обратимых и необратимых процессов, структурная и функциональная организация потоков энергии, эндогенные источники энергии и информации, память, концентрация внимания, доминанта, торможение, асимметрия, способность регулировать гомеостазис и степень «открытости» организма как неустойчивой термодинамической системы во взаимодействии с окружающей средой. Эти свойства позволяют поддерживать достаточно низкую скорость роста энтропии, а также быть относительно независимыми от окружающей среды. Можно предположить, что термодинамическая «пластичность» и разнообразие путей «обхода» запрета второго начала термодинамики живыми организмами являются весомыми факторами, определяющими специфику живого (Чернышева, Ноздрачев, 2006). Кроме того, процессы жизнеобеспечения, связанные с делением, ростом и дифференцировкой клеток, метаморфозами и регенерацией, движением и поведением, не приводят к «тепловой смерти», но сохраняются в течение жизни и могут передаваться генетически благодаря названным «антиэнтропийным механизмам» (термин Ю. А. Романова, 2000).
Общеизвестно, что живые организмы как открытые термо-динамические системы обмениваются с окружающей средой материей, энергией, информацией и, добавим, временем. Последнее согласуется с тезисом о существовании времени только для открытых систем (Левич, 2013). Для успешности такого обмена необходимым условием является способность биосистемы создавать материю, генерировать энергию, информацию и время. Это подтверждают способность к образованию молекул веществ в процессах синтеза, метаболизма нутриентов поглощаемой пищи и катаболизма синтезированных веществ, а также выделение энергии в реакциях метаболизма, дефосфорилирования макроэргов (АТФ, ГТФ, КФ и др.) и других молекул или же их депротонирования и т. д. В частности, известно, что в процессах генеза и процессинга информации в нервной системе усиление активности Na,K-ATФазы клеточной мембраны нейрона на фазе следовой гиперполяризации потенциала действия приводит к восстановлению асимметрии концентрации ионов Na+ и K+ и потенциала покоя мембраны, а реаптейк транспортерами молекул нейромедиатора на уровне пресинаптической мембраны осуществляется на градиенте Н+ или Na+. Известно, что процессы сокращения и расслабления скелетных мышц при поддержании позы или двигательной активности также осуществляются при участии Na,K-ATPазы и Са,Mg-ATPазы, способных присоединять молекулы АТФ, дефосфорилировать их, а выделившуюся энергию частично использовать на перенос ионов через мембраны против градиента их концентраций. Каждый из этих процессов характеризуют временные параметры (латентность, длительность, скорость), что позволяет говорить о них как о временных процессах.
Постулируем взаимосвязь биологического времени с информацией, метаболизмом и энергией. Для формализации такой взаимосвязи рассмотрим особенности взаимодействия информации и времени, а также метаболизма и времени.
1.2. Информация и биологическое время
Известный тезис Аристотеля «Время является мерой движения (изменения)» (Аристотель, 1937) применительно к живым организмам может быть переформулирован как «время есть мера изменения информации» В пользу этого тезиса свидетельствует ряд исследований, в частности, работа R.E. Hicks и соавторов (Hicks et al., 1976), в которой авторы рассматривают проспективные и ретроспективные суждения о времени как функцию от объема полученной информации. О схожей закономерности, связывающей время и информацию в биосистемах писал М. И. Сетров (1974).
Проанализируем в этом аспекте два основные определения: информация как сообщение/сигнал о чем-либо и информация как негэнтропия (Шредингер, 2002; Бриллюэн, 2006). Другие определения условно можно считать по смыслу близкими первому или второму из них, дополняющими характеристику свойств/функций информации. Многочисленные данные из различных областей биологии свидетельствуют о справедливости для биосистем обоих определений, а также об одновременном взаимосвязанном генезе информации и эндогенного времени на разных структурных уровнях организма (Чернышева, 2011). Рассмотрим эти положения более конкретно.