
Заболевания и травмы периферической нервной системы
Под влиянием естественной или вызванной внешними факторами стимуляции аксона происходит нарушение селективной проницаемости клеточной мембраны, что способствует проникновению ионов натрия в клетку и редукции потенциала покоя. Если мембранный потенциал снижается (деполяризуется) до критического уровня (30 – 50 мВ), то возникает потенциал действия, и импульс начинает распространяться вдоль клеточной мембраны как волна деполяризации. Важно отметить, что в немиелинизированных волокнах скорость распространения импульса прямо пропорциональна диаметру аксона, и возбуждение длительно прямолинейно захватывает соседствующие мембраны (рис. 1.9, 1.10).
Проведение импульса в миелинизированных волокнах совершается «сальтаторно», т. е. как бы скачкообразно: импульс или волна деполяризации мембраны скользит от одного перехвата Ранвье до другого, и так далее (цветная вклейка, рис. 1.11). Миелин действует как изолятор и предупреждает возбуждение мембраны клетки аксона, за исключением промежутков на уровне перехватов (узлов) Ранвье. Нарастание проницаемости возбужденной мембраны этого узла для ионов натрия вызывает ионные потоки, которые и являются источником возбуждения в области следующего перехвата Ранвье.
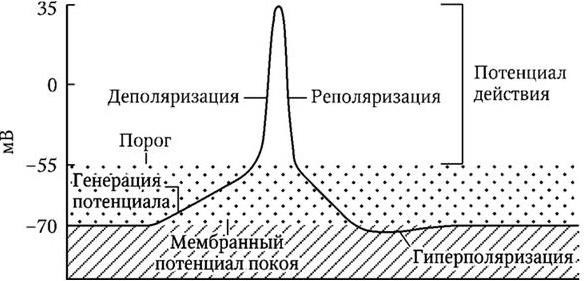
Рис. 1.9. Развитие пика составного потенциала действия нервного ствола
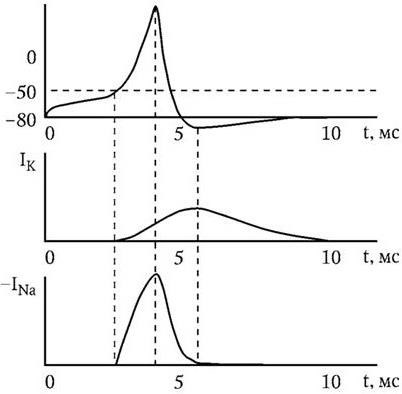
Рис. 1.10. Зависимость ионных токов и мембранного потенциала от времени
Таким образом, в миелинизированных волокнах скорость проведения импульса зависит не только от диаметра аксона и толщины миелинового футляра, но и от дистанции между узлами Ранвье (от «интернодальной» длины).
Физиология синаптической передачи. Информация, передаваемая по нерву, распространяется не только стереотипными электрическими сигналами, но и с помощью химических передатчиков нервного возбуждения – медиаторов или трансмиттеров, высвобождаемых в местах соединения клеток – синапсах. Синапсы – специализированные контакты, через которые осуществляется поляризованная, опосредованная химически, передача из нейрона возбуждающих или тормозящих влияний на другой клеточный элемент. Различают аксодендритические синапсы – аксон оканчивается на дендрите, аксосоматические синапсы – образуется контакт между аксоном и телом нейрона, аксо-аксональные синапсы – контакт устанавливается между аксонами. В окончаниях периферических нервов также имеются нервно-мышечные (мионевральные) синапсы. Примерно половина всей поверхности тела нейрона и почти вся поверхность его дендритов покрыты синаптическими контактами от других нейронов (рис. 1.12; цветная вклейка, рис. 1.13).
Рис. 1.12. Аксосоматические синапсы
В дистальной части нервное волокно лишено миелина, образуя терминальную арборизацию (телодендрон) и пресинаптический терминальный элемент. Этот элемент морфологически характеризуется расширением окончания аксона, что напоминает булаву и нередко именуется «пресинаптический мешок», «терминальная бляшка», «синаптический узелок». Под микроскопом в этой булаве можно увидеть различных размеров – около 0,05 мкм (500 А) – гранулярные пузырьки или синаптические везикулы, содержащие медиаторы (ацетилхолин, катехоламины, пептидные гормоны и др.). Под терминальной бляшкой лежит синаптическая щель размерами 0,2 – 0,5 мкм в поперечнике, в которую из везикул поступают кванты медиатора. Затем следует субсинаптическая (постсинаптическая) мембрана, воздействуя на которую химический передатчик вызывает изменения электрического потенциала в подлежащих клеточных элементах (цветная вклейка, рис. 1.14).
Порядок синаптической передачи (цветная вклейка, рис. 1.15). 1. Импульс подходит к пресинаптической части волокна. Открываются кальциевые каналы, ионы Ca2+ поступают в пресинаптическую цитоплазму.
2. В пресинаптической цитоплазме постоянно находится большое количество синаптических пузырьков – образований, содержащих молекулы вещества-медиатора. Вследствие появления ионов Ca2+ в пресинаптической цитоплазме пузырьки подходят к мембране и выделяют медиатор в синаптическую щель. Попавший в щель медиатор диффундирует на постсинаптическую сторону щели. Этот процесс идет около 0,5 мс и вносит существенный вклад в задержку передачи.
3. Молекулы медиатора улавливаются рецепторами на постсинаптической стороне.
4. Попадание ацетилхолина на рецептор увеличивает проводимость канала в обоих направлениях для ионов Na+ и K+ в равной степени. Открытие каналов на постсинаптической стороне вызывает поток ионов натрия внутрь, а калия – наружу. Возникший ионный ток возбуждает нервный импульс, который распространяется дальше по постсинаптическому волокну.
5. Происходит обратный захват медиатора пресинаптическим окончанием.
Типы нервных волокон. Согласно А. Paintal (1973), периферические нервные волокна по характеру строения, проведению возбуждения, длительности фаз потенциала действия подразделяются на следующие классы:
Класс А: 1. Миелинизированные волокна, афферентные, сенсорные.
Группа I. Волокна диаметром более 20 мкм, со скоростью проведения импульса до 100 м/с. Волокна этой группы несут импульсы от рецепторов мышц (мышечных веретен, интрафузальных мышечных волокон) и рецепторов сухожилий.
Группа II. Волокна диаметром от 5 до 15 мкм, со скоростью проведения импульсов от 20 до 90 м/с. Эти волокна несут импульсы от механорецепторов и вторичных окончаний на мышечных веретенах интрафузальных мышечных волокон.
Группа III. Волокна диаметром от 1 до 7 мкм, со скоростью проведения импульса от 12 до 30 м/с. Их функциями являются болевая рецепция, а также иннервация волосяных рецепторов и сосудов.
2. Миелинизированные волокна, эфферентные, двигательные.
Альфа-волокна. Более 17 мкм в диаметре, скорость проведения импульса от 50 до 100 м/с. Они иннервируют экстрафузальные поперечно-полосатые мышечные волокна, преимущественно стимулируя быстрые сокращения мышц (мышечные волокна 2-го типа) и крайне незначительно – медленные сокращения (мышц 1-го типа).
Бета-волокна. В отличие от альфа-волокон иннервируют мышечные волокна 1-го типа (медленные и тонические сокращения мышц) и частично интрафузальные волокна мышечного веретена.
Гамма-волокна. Размером 2 – 10 мкм в диаметре, скорость проведения импульса 10 – 45 м/с, иннервируют только интрафузальные волокна, т. е. мышечное веретено, тем самым участвуя в спинальной саморегуляции мышечного тонуса и движений (кольцевая связь гамма-петли).
Класс В – миелинизированные волокна, преганглионарные, вегетативные.
Небольшие нервные волокна, около 3 мкм в диаметре, со скоростью проведения импульса от 3 до 15 м/с.
Класс С – немиелинизированные волокна.
Волокна диаметром от 0,2 до 1,5 мкм, со скоростью проведения импульса от 0,3 до 1,6 м/с. Этот класс волокон состоит из постганглионарных вегетативных и эфферентных волокон, преимущественно воспринимающих (проводящих) болевые импульсы.
Периферическая нервная система. Анатомическое образование, включающее комплекс нервных волокон и соединительнотканных оболочек, получило название нервного ствола. Нервные стволы связывают ЦНС с соматическими и висцеральнымиорганами, а также висцеральные органы между собой, создавая таким образом основу для местной рефлекторной регуляции. В связи с этим нервы условно подразделяются на соматические и висцеральные (вегетативные, автономные). По топографическому принципу выделяют спинномозговой и краниальный отделы ПНС.
Спинномозговой отдел периферической нервной системы включает анатомические образования, связанные со спинномозговыми нервами:
– передние и задние корешки спинномозговых нервов (radix ventralis et radix dorsalis). Спинномозговые нервы образуются в результате слияния чувствительных волокон (отростков клеток спинномозговых узлов, формирующих дорсальные чувствительные корешки) с пучками двигательных (эфферентных) волокон, являющихся отростками клеток передних рогов спинного мозга и составляющих передние (вентральные) двигательные корешки. У человека имеется 31 пара спинномозговых нервов: 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1 копчиковая;
– ствол спиномозгового нерва (truncus n. spinalis);
– спинномозговой узел (чувствительный) (ganglion spinale);
– ветви спиномозгового нерва: передние (вентральные), задние (дорсальные), соединительные, менингеальные. Вентральные иннервируют кожу, мышцы конечностей и передней поверхности туловища, дорсальные иннервируют кожу и мышцы задней поверхности туловища, соединительные содержат симпатические преганглионарные волокна, следующие к узлам симпатического ствола, менингеальные направляются к твердой оболочке спинного мозга. Передние ветви грудного отдела дают начало межреберным (nn. intercostales) и подреберному (n. subcostalis) нервам;
– нервные сплетения: шейное, плечевое, поясничное, крестцовое, копчиковое, из них берут начало периферические кожные и мышечные нервы, в формировании которых обычно принимают участие несколько сегментов спинного мозга;
– регионарные (органные) нервы и их ветви;
– нервные окончания (рецепторы и эффекторы).
Краниальный отдел периферической нервной системы представлен черепными нервами и их узлами, регионарными (органными) нервами и их ветвями, нервными окончаниями. Черепные нервы (12 пар) начинаются в головном мозге от соответствующих ядер, покидают полость черепа (от них в области головы и шеи ответвляется блуждающий нерв) и следуют в органы грудной и брюшной полостей. В них проходят аксоны разной функциональной специализации. Соматическая чувствительность осуществляется II, V, VIII парами, соматодвигательные функции обеспечиваются III, IV, V, VI, VII, IX, X, XI, XII парами, висцерочувствительные – I, V, VII, IX, X парами и, наконец, висцеродвигательные функции – VII, IX, X парами.
Автономная нервная система. К периферическому отделу автономной нервной системы относят:
– вегетативные (автономные) нервы, ветви и нервные волокна, выходящие из головного и спинного мозга;
– вегетативные (автономные, висцеральные) сплетения;
– узлы вегетативных (автономных, висцеральных) сплетений;
– симпатический ствол (правый и левый) с его узлами, межузловыми и соединительными ветвями и симпатическими нервами;
– концевые узлы (ganglia terminalia) парасимпатической части вегетативной нервной системы.
На основании топографии вегетативных ядер и узлов, различий в длине первого и второго нейронов эфферентного пути, а также особенностей функции вегетативная нервная система подразделяется на две части: симпатическую и парасимпатическую (рис. 1.16).
Парасимпатическая нервная система. Парасимпатическая нервная система состоит из краниальной и сакральной частей (цветная вклейка, рис. 1.17). Тела клеток преганглионарных парасимпатических нейронов краниальной части АНС располагаются в ядрах черепных нервов в стволовой части мозга. Их аксоны проходят к периферическим ганглиям в составе глазодвигательного, лицевого, языкоглоточного и блуждающего нервов. Длинные преганглионарные волокна, выходящие с этими нервами, образуют синапсы с короткими постганглионарными волокнами, исходящими из ресничного, клиновидно-небного и слухового ганглиев. Наиболее длинные преганглионарные волокна блуждающего нерва образуют синапсы с интрамуральными ганглиями в сердце, легких и в желудочно-кишечном тракте (рис. 1.18).
Тела клеток преганглионарных нейронов крестцовой части парасимпатической нервной системы локализуются в боковом роге спинного мозга на уровне II – IV крестцовых сегментов. Их аксоны выходят в составе передних корешков спинномозговых нервов, а затем формируются в нервы, образующие тазовое сплетение.
Окончания аксонов находятся в терминальных ганглиях таза, в висцеральных сплетениях или в интрамуральных ганглиях мочевого пузыря, в нисходящей, сигмовидной и в прямой кишке, а также в половых органах.
Симпатическая нервная система. Тела клеток преганглионарных нейронов находятся в боковом роге серого вещества спинного мозга в грудопоясничном отделе на уровне от ТhI до LII. Аксоны этих преганглионарных нейронов выходят в составе передних корешков спинномозговых нервов или в виде коммуникационных веточек, проникая в паравертебральные ганглии симпатической цепочки (цветная вклейка, рис. 1.19). Дальнейший их путь различен: 1) синапсы в ганглиях симпатической цепочки; 2) прохождение вверх или вниз по симпатической цепочке, прежде чем образовывать синапсы; 3) прохождение непосредственно к превертебральным ганглиям или к сплетениям (брюшное, верхнее мезентериальное и др.).
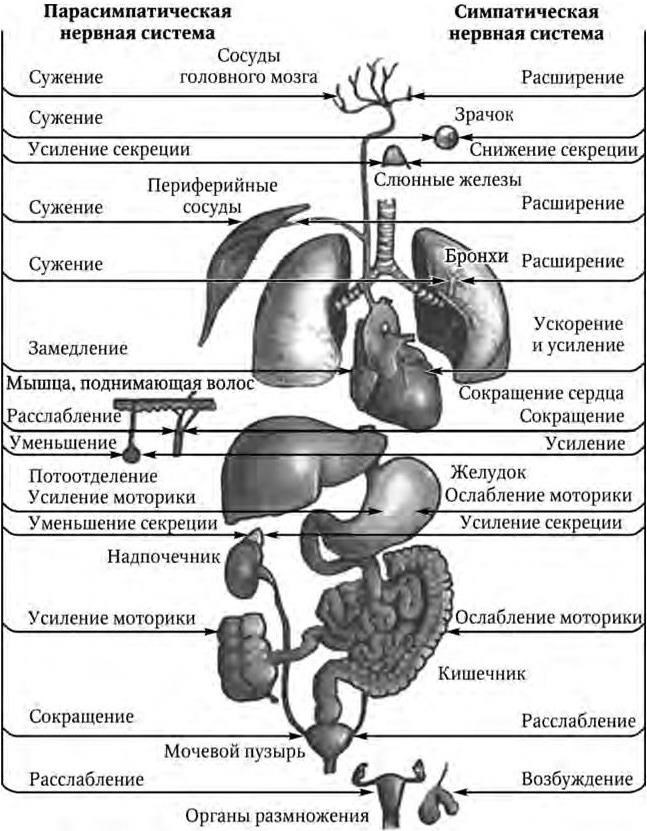
Рис. 1.16. Автономная нервная система
Рис. 1.18. Интрамуральный ганглий
Паравертебральные ганглии расположены попарно и проходят двумя рядами по обеим сторонам позвоночника от II шейного позвонка до копчика. Восходящие и нисходящие нервные волокна соединяют между собой соседние ганглии, формируя таким образом цепочку, или ствол. Шейные узлы расположены кпереди от оснований соответствующих поперечных отростков позвонков. Поясничные ганглии располагаются на переднебоковой поверхности соответствующих позвонков. Превертебральные ганглии лежат дистальнее симпатической цепочки, несколько проксимальнее их окончаний в органах.
Глава 2
ПАТОГЕНЕЗ ТРАВМАТИЧЕСКИХ ПОРАЖЕНИЙ И ЗАБОЛЕВАНИЙ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
2.1. Патогенез травматических невропатий и плексопатий
2.1.1. Механические факторы, вызывающие повреждения периферической нервной системы
Основной причиной двигательных, чувствительных и вегетативно-трофических расстройств при травмах ПНС являются прекращение аксонального транспорта и блокада невральной проводимости возбуждения, вызванные нарушением анатомической целостности осевых цилиндров (аксонов) на макро- или микроскопическом уровне.
Знание механических свойств различных участков ПНС позволяет ускорить и улучшить топическую и патогенетическую диагностику при конкретных видах травматических поражений. Поскольку нервный ствол является гетерогенным полуэластичным образованием, имеющим прочный соединительнотканный каркас, он более устойчив к компрессии, чем к тракции (эластичный лимит составляет 20 % от исходной длины). Спинномозговые корешки, лишенные эпиневральной и периневральной оболочек, являются самыми уязвимыми элементами ПНС.
Кроме того, периферические нервы и сплетения не только представляют собой органную, гетерогенную в морфофункциональном отношении систему, состоящую из нервной, глиальной и соединительной ткани и характеризующуюся сложными межтканевыми взаимоотношениями, но и являются связующим звеном между периферическим аппаратом (кожа, мышцы) и командным центром (ЦНС).
Закрытая либо открытая травма нерва (сплетения) в очень редких случаях является изолированной; в абсолютном большинстве наблюдений нервные образования страдают вместе с другими тканями конечности, что накладывает существенный отпечаток на характер развития неврологического симптомокомплекса.
Открытые повреждения нервов, при которых нарушается целостность эпиневрия, вызываются непосредственным действием ранящего агента на структурное образование ПНС и подразделяются на колотые, резаные, рубленые, огнестрельные и т. д.
Патогенез современных огнестрельных повреждений ПНС имеет существенные особенности, так как современное вооружение по характеру и мощности значительно отличается от вооружения, применяемого в годы Второй мировой войны. Широкое применение в международных и внутренних конфликтах, а также в террористических акциях находят стрелковое малокалиберное скорострельное оружие с высокой начальной скоростью полета пули с малой устойчивостью, а также мощные взрывные устройства, что значительно повысило эффективность и плотность огня, а также поражающие свойства.
При огнестрельных ранениях конечностей нервный ствол и/или сплетение повреждается либо непосредственно ранящим снарядом (напрямую или за счет бокового удара раневой пульсирующей полости), либо вовлекается в посттравматический рубцово-спаечный процесс. Сотрясение, ушиб нерва, миллиарные кровоизлияния в нем, периневральные спайки являются более частыми причинами параличей после огнестрельных ранений конечностей, чем невротмезис.
Установлено, что ударная волна (раневая пульсирующая полость), распространяясь преимущественно по ходу рыхлой клетчатки между фасциями, способна разорвать спинномозговые корешки, нервные волокна и сосуды внутри нерва в отдалении от раны, хотя сам нервный ствол может казаться внешне неповрежденным. Поражающий эффект раневой пульсирующей полости обнаружен в эксперименте даже в головном мозге при огнестрельном ранении бедра животного.
Кроме того, при опосредованных повреждениях ПНС субэпиневральные кровоизлияния иногда возникают на некотором расстоянии от линии полета пули, причем не только сразу, но и на вторые сутки после ранения, а гематомы, даже незначительные, вызывают периаксональные изменения нервных волокон и рассасываются спустя 2 – 3 месяца, оставляя после себя микроскопические внутриствольные рубцы.
По данным В. Л. Попова [и др.] (1990), изменения в нерве в зоне 2 – 4 см от оси раневого канала характеризуются мощными очаговыми кровоизлияниями в субпериневральное пространство и в эндоневрий, резким отеком и воспалительными инфильтратами во всех его тканях. Миелиновые нервные волокна резко гипертрофированы, часть из них находится в состоянии валлеровской дегенерации, миелиновые сегменты большинства нервных волокон содержат очень большое число вакуолей. На удалении 5 – 6 см отечные явления в нерве ослабевают, реже встречаются воспалительные элементы и кровоизлияния, нервные волокна приобретают близкий к норме вид.
Механизмы опосредованного поражения нервов и сплетений при огнестрельных повреждениях отражены в литературе недостаточно. Н. П. Бисенков, М. И. Попович (1980) полагают, что при огнестрельных ранениях в результате пульсации временной раневой полости нервы, расположенные в стороне от раневого канала, испытывают неоднократное сжатие и кратковременное, но сильное растяжение, которое и приводит к повреждению различных элементов нервной системы, в первую очередь, периферической.
Более осторожное заключение о тракционном механизме огнестрельных повреждений нервов конечностей делается в исследованиях К. А. Григоровича (1981). При сравнении огнестрельных и неогнестрельных повреждений нервов автор отметил, что ранения высокоскоростными или крупнокалиберными ранящими снарядами часто приводят к протяженным изменениям нервов и требуют относительно больших трансплантатов с худшими результатами лечения.
Обобщив данные литературы и результаты собственных исследований, мы разработали оригинальную схему патогенеза огнестрельных поражений ПНС (схема 2.1).
Минно-взрывные ранения и травмы ПНС – это особый вид огнестрельных поражений организма, так как возникают в результате действия не одного травмирующего агента (например, осколок или вторичные ранящие снаряды, ударные волны высокой энергии), а комплекса поражающих факторов взрыва минного боеприпаса, особенно при контактном механизме подрыва человека или технического средства. Минно-взрывная травма представляет собой классический пример огнестрельной, но сочетанной и множественной травмы – политравмы. В основе ее патогенеза лежит многофакторный характер поражений ПНС.
При закрытых неогнестрельных травматических поражениях нервов и сплетений выделяют следующие основные причины нарушения невральной проводимости и аксоплазматического тока:
1) поражение длинных сегментов нерва в связи с его растяжением вследствие смещения конечности;
2) сдавление, размозжение или контузия нерва, обычно возникающие при падении, компрессии конечности при дорожных происшествиях, иногда причиняемые ударом тупыми предметами;
3) сдавление нерва за счет отека под тугой манжетой или повязкой или его контузия;
4) ушиб, растяжение нерва (или одновременно оба вида повреждений) или последующее включение нерва в костную мозоль при сращении переломов;
5) повреждения, вызванные инъекциями;
6) «компартмент-синдром» – резкое повышение внутритканевого давления в костно-фасциальных пространствах конечностей, наблюдающийся в 45 % случаев переломов костей. Возникающая при этом ишемия может без лечебных вмешательств привести к необратимым изменениям тканей, расположенных в костно-фасциальных футлярах.
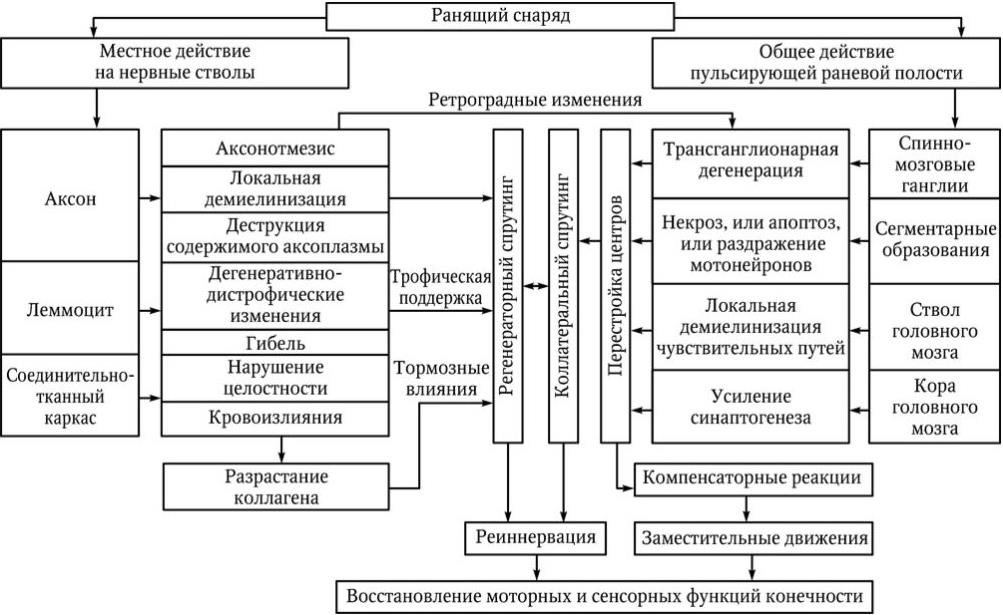
Схема 2.1. Патогенез огнестрельных невропатий и плексопатий
Если различные виды травматических поражений нервов конечностей и сплетений освещены в литературе в достаточной степени, то механизмы повреждающего действия ранящих факторов на нервные стволы и сплетения разные авторы до сих пор объясняют по-разному. Ряд исследователей считают, что при травмах конечностей нервы повреждаются в результате тракции, трения и компрессии внутренними или внешними силами, причем нервы устойчивы к большим степеням растяжения и сдавления при постепенном развитии деформации. Другие авторы полагают, что подавляющее большинство закрытых повреждений нервов по механизму являются тракционными, а ушиб нерва как самостоятельная нозологическая форма без момента тракции наблюдается сравнительно редко.
В свою очередь, компрессионные травмы нервов и сплетений могут быть частичными или полными, локализованными или протяженными по длине нервов. К настоящему времени выполнено много экспериментальных исследований, посвященных изучению изменений в нервах при их острой и хронической компрессии. В этих работах показано, что даже кратковременная, но сильная компрессия нерва приводит к блокаде невральной проводимости за счет локальной демиелинизации в месте сдавления, дегенерации части нервных волокон с последующей прогрессирующей дистальной атрофией нерва. В то же время умеренная тракция нервных стволов (400 г на седалищный нерв кролика) приводила к гиперемии, геморрагиям в эпиневрий и периневрий, разрыву соединительнотканных волокон, нарушению целостности части аксонов с последующей дегенерацией их дистальных участков.
При выраженной тракции, вследствие уменьшения поперечного сечения нервных стволов, повышается внутрифасцикулярное давление, что, в свою очередь, вызывает компрессионную деформацию их содержимого, в том числе кровеносных сосудов, формируя ишемию невральных тканей. Лимитом эластичности нервов является 20 % от длины, а порогом разрыва пучков – 30 %.
Таким образом, очевидна целесообразность выделения в отдельные группы механических факторов огнестрельных и неогнестрельных повреждений ПНС, вследствие принципиального различия объемов поражающего действия.
С другой стороны, механизмы огнестрельных и неогнестрельных повреждений нервных волокон во многом сходны и в конечном счете могут быть сведены к компрессии, тракции и ишемии (их сочетанию). Однако данное предположение нуждается в тщательной проверке на предмет выяснения соотношения значимости этих механизмов при различных видах травматических невропатий и плексопатий.
Кроме того, несмотря на системный подход при оценке поражающего действия механических факторов на ПНС, для лучшего понимания патологических процессов представляется целесообразным выделить различные уровни в структуре нейромоторного аппарата, обеспечивающего реализацию жизненно важных приспособительных функций организма.
2.1.2. Морфологическая и функциональная характеристика травматических невропатий и плексопатий
Морфологические и функциональные реакции нервов и сплетений на механическую травму определяются уровнем, характером и тяжестью полученного повреждения, причем различные компоненты нервов (соединительнотканные образования, нервные волокна, собственная сосудистая сеть) по-разному реагируют на повреждающие воздействия.
Для травм с полным или частичным нарушением анатомической целостности нерва или сплетения (невротмезис) характерна валлеровская дегенерация дистального отрезка нервного ствола с разрастанием коллагена в проксимальных участках прерванных нервных пучков и нарушением гистогематического барьера между кровью и нервом.
Особенности валлеровской дегенерации к настоящему времени изучены достаточно глубоко. В частности, показано, что пусковым механизмом может быть большое поступление ионов кальция в поврежденный участок нерва. Изменения мякотной оболочки носят более выраженный и продолжительный характер, чем аксона. В распаде миелиновых волокон различают несколько стадий.