


Основы нейропсихологии. Теория и практика
Кора мозга делится на следующие доли (рис. 34):
• затылочная (зрительная) доля;
• теменная (тактильная) доля;
• височная (слуховая) доля;
• лобная (управляющая, регулирующая) доля.
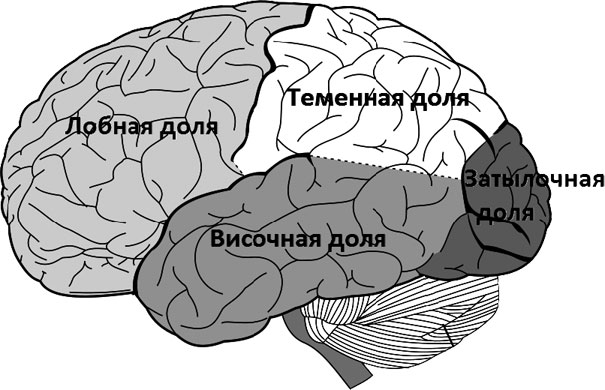
Рис. 34. Доли коры мозга
Затылочная, теменная и височная доли имеют соответствующую анализаторную отнесенность. В нейропсихологии ее принято обозначать как модальную специфичность. Благодаря им осуществляются разные психические функции. Вкусовые и обонятельные отделы расположены на медиальной (внутренней) поверхности височной доли. Их роль в осуществлении когнитивных функций у современного человека перестала быть ведущей, то есть уступает по функциональной значимости ролям остальных долей мозга.
Лобная доля не имеет модальной специфичности, но играет главенствующую роль в осуществлении высшей нервной деятельности человека. Она занимает обширную площадь (более половины коры) и отвечает за все мозговые процессы.
Во многих публикациях по неврологии и нейрофизиологии отмечается, что сложнейшая мозговая деятельность обеспечивается, в сущности, простыми средствами. Некоторые из авторов отмечают, что эта простота отражает универсальный закон «достижения большой сложности через многократные преобразования простых элементов» (Э. Голдберг). Так, множество слов в языке складывается из ограниченного числа звуков речи и букв алфавита, бесчисленные музыкальные мелодии – из малого числа нот, генетические коды миллионов людей обеспечиваются конечным числом генов и т. д.
Глава 2. Поля коры мозга
Понятие полей коры мозга и их функциональной иерархии
Представления о дифференциации коры головного мозга на три основных, различных по функциональной иерархии вида полей: первичные, вторичные и третичные – являются чрезвычайно важными для понимания того, как организована психика человека в целом.
Первичные поля – это «корковые концы анализаторов», функционирующие от природы, врожденно. Первичные поля имеют четкую отнесенность к тому или иному анализатору.
Элементарными являются первичные поля, более непростыми по строению и функционированию – вторичные и, наконец, максимально сложными по этим признакам – третичные поля (рис. 35).
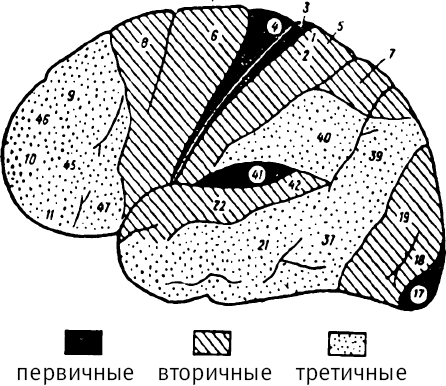
Рис. 35. Функциональная иерархия полей коры мозга
Первичные поля слуховых анализаторов располагаются преимущественно на внутренней поверхности височных долей мозга, кинестетического (чувствительного в целом) – вблизи от центральной (Ролландовой) борозды, в теменной доле. Первичные чувствительные поля являются проекционными в отношении определенных частей тела: верхние отделы принимают чувствительные сигналы (ощущения) от нижних конечностей (ног), средние отделы обрабатывают ощущения от верхних конечностей (рук), а нижние отделы – от лица, включая отделы речевого аппарата (язык, губы, гортань, диафрагму). Кроме того, нижние отделы теменной проекционной зоны принимают ощущения от некоторых внутренних органов.
Первичные поля, расположенные на мозговой территории до центральной извилины (переднем блоке мозга), настроены на подготовку и исполнение двигательных актов. Они также являются проекционными, но уже в отношении не чувствительных (кинестетических – сенсорных), а двигательных (моторных) функций. На уникальных рисунках, сделанных известным исследователем работы мозга У. Пенфильдом (W. G. Penfield), видно, что значимость разных частей тела не совпадает с их размерами, а определяется той ролью, которую они играют в осуществлении психических функций как при восприятии объектов внешнего мира, так и при воспроизведении различных действий (рис. 36).
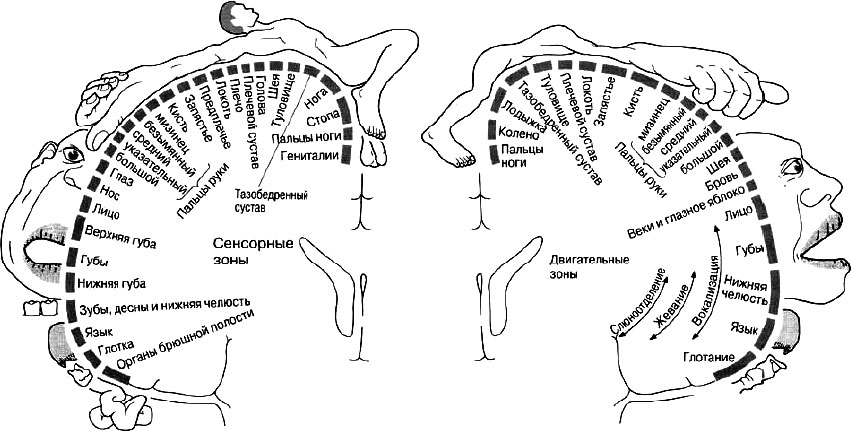
Рис. 36. Функциональное представительство в коре мозга частей тела человека
Нервные клетки первичной коры мозга в самом раннем онтогенезе функционируют изолированно друг от друга, подобно отдельным мирам в космосе. Допустим, ребенок узнает голос матери, но не узнает ее лицо, если она молчит. Особенно часто такое разобщение слуховых и зрительных впечатлений на уровне ощущений наблюдается в отношении отца, которого младенцы видят реже, чем мать. В литературе описаны случаи, когда ребенок, увидев склоненное над ним отцовское лицо, начинает громко испуганно плакать, пока взрослый не заговорит. Постепенно между первичными полями коры мозга прокладываются смысловые, а точнее информационные связи (ассоциации). Благодаря им накапливается опыт ощущений, то есть появляются элементарные знания о действительности. Например, ребенок видит погремушку и знает, что она будет «греметь», если ее потрясти.
Опыт, накопленный взаимодействующими между собой первичными полями, служит базой, отправным моментом для функциональной активизации вторичных полей коры вместе с третичными полями, о которых речь пойдет далее. И те и другие имеют непосредственное отношение к реализации ВПФ.
Выделяются вторичные поля слуховой, тактильной и зрительной коры, а в переднем – премоторной. Функционально все три вида полей коры соотносятся между сбой вертикально: функции первичных, над ними надстраиваются функции вторичных, а над вторичными – третичных. Однако анатомически они располагаются не друг над другом, а по горизонтали: первичные поля (I) близко к ядру зон, вторичные (II) – в ее срединных отделах, а третичные (III) – на периферии (рис. 37).
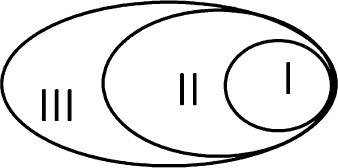
Рис. 37. Схема функциональной иерархии полей коры мозга
Первичные поля составляют ядро той или иной анализаторной зоны. Вторичные поля сдвинуты к периферии зоны, а третичные – еще дальше. Пропорциональны близости к ядру и размеры разных по иерархии полей: первичные занимают маленькую площадь, вторичные – среднюю, а третичные – самые большие. Вследствие этого поля накладываются друг на друга, образуя так называемые зоны «перекрытия». К ним относится, например, самая важная в рамках высшей деятельности человека зона ТРО – височно-теменно-затылочная: Temporalis – Parietalis – Occipitalis. Три первые буквы этих латинских обозначений и составляют аббревиатуру TPO.
Первичные поля однородны по клеточному составу. Обонятельные поля содержат только обонятельные нервные клетки, слуховые – только слуховые и т. п. Несмотря на универсальность физиологических и биохимических механизмов, обеспечивающих работу мозга, его различные отделы функционируют по-разному, то есть имеют различную функциональную специализацию, представляя разные модальности.
Вторичные поля менее однородны. В состав клеток преобладающей модальности вкраплены клетки других модальностей. Поэтому вторичные поля, хоть имодально-специфичны, подобно первичным, но являются менее «жесткими» по этому признаку. Третичные поля, будучи зонами перекрытия, содержат не только клетки разных анализаторов, но и их целые зоны. Благодаря их функционированию реализуются наиболее сложные виды деятельности человека, и в частности речевая.
Вторичные и третичные поля коры отличаются разницей в функционировании в зависимости от латерализации, то есть расположенности в том или другом полушарии мозга. Например, обе височных доли, относясь к одной и той же, а именно слуховой модальности, выполняют разную «работу». Височная доля правого полушария, например, ответственна за обработку неречевых шумов (издаваемых природой, включая звуки животных, предметов, в том числе музыкальных инструментов и саму музыку, которую можно считать высшим видом неречевого шума). Височная доля левого полушария осуществляет обработку речевых сигналов. Помимо различий в специализации височных долей мозга, относящихся к разным полушариям, здесь можно усмотреть и столь характерный для природы принцип «защиты» наиболее важных функций, и тем более такой важной и необходимой любому человеку, как речь.
Разрушение первичных полей невосполнимо, то есть утерянные физический слух, зрение, обоняние и пр. не восстанавливаются. Функции поврежденных вторичных полей подлежат компенсации за счет подключения других, «здоровых» систем мозга и перестройки способа их деятельности. Нарушения функций пострадавших третичных полей компенсируются относительно легко за счет их полимодальности, позволяющей опираться на мощную систему ассоциаций, хранящихся в каждом из полей и между ними.
Различия в функциональной специфике первичных, вторичных и третичных полей обусловливают и различия в их способности заменять друг друга (компенсировать) в случае патологии.
Необходимо, однако, помнить, что и в этом случае важное значение имеют возрастные пороги и время, когда начаты восстановительные мероприятия. Наиболее благоприятны ранний возраст и раннее начало коррекционно-восстановительного обучения.
Слои коры мозга
Несмотря на то что кора мозга (серое вещество) имеет малую толщину всего 1,3–4,5 мм, она состоит из 6 слоев (рис. 38). Вызывает немалое удивление то, каким образом в ней располагается множество различно функционирующих нейронов, составляющих серое вещество, проводниковые системы, глиальный слой. Серое вещество покрывает кору подобно большой складчатой салфетке. Этот слой отвечает за сенсорное восприятие, генерацию моторных команд, пространственное ориентирование, сознательное и абстрактное мышление, речь и воображение. Увеличение объема неокортекса регулируется сроками нейрогенеза (образования нервной ткани), которые, разумеется, находятся под контролем ДНК.
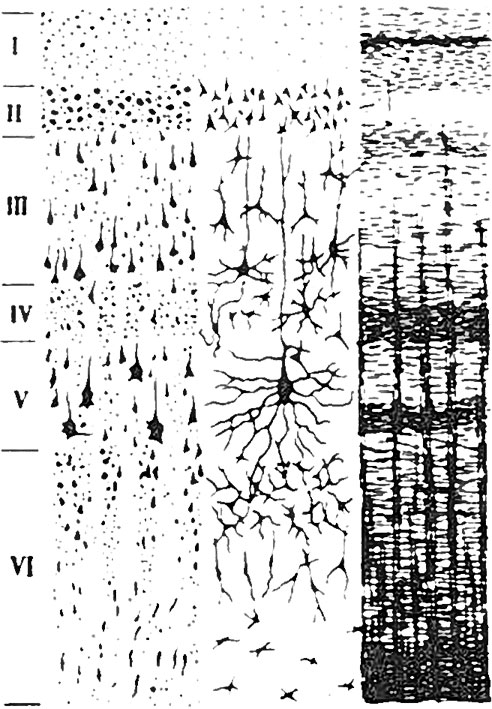
Рис. 38. Слои коры мозга
Между слоями коры расположена глия (совокупность вспомогательных клеток нервной ткани), которая выполняет, по существу, роль клея, связывающего многочисленные нейроны коры мозга между собой. Тем самым глия участвует в межзональных связях с когнитивной сутью. Микроскопические размеры отделов, составляющих кору мозга, потребовали изобретения невероятно мощных технических средств для их изучения. В настоящее время исследователи в области нейронаук вооружены приборами, позволяющими увидеть то, о существовании чего не так давно даже не знали. В результате перестало быть тайной, что кора мозга представлена несколькими слоями.
I. Молекулярный. Клеток в нем мало, в основном горизонтальные волокна восходящих аксонов, в том числе неспецифические афферентные, идущие от таламуса.
II. Наружный зернистый. Состоит из звездчатых и мелких пирамидальных клеток, аксоны которых заканчиваются в 3, 5 и 6-м слоях, то есть участвует в соединении различных слоев коры.
III. Наружных пирамид. Этот слой имеет два подслоя. Внешний состоит из более мелких клеток, которые осуществляют связь с соседними участками коры, особенно хорошо он развит в зрительной коре. Внутренний подслой содержит более крупные клетки, которые участвуют в образовании коммиссуральных связей (между двумя полушариями).
IV. Внутренний зернистый. Включает клетки зернистые, звездчатые и мелких пирамид. Их апикальные дендриты поднимаются в 1-й слой коры, а базальные (от основания клетки) в 6-й слой коры, таким образом участвуют в осуществлении межкорковой связи.
V. Ганглиозный. Основу этого слоя составляют гигантские пирамиды (клетки Беца). Их апикальный дендрит простирается до 1-го слоя, базальные дендриты идут параллельно поверхности коры, а аксоны образуют проекционные пути к базальным ядрам, стволу и спинному мозгу.
VI. Полиморфный. В нем присутствуют клетки различной формы, но преимущественно – веретенообразные. Их аксоны идут вверх, но в большей мере вниз и образуют ассоциативные и проекционные пути, переходящие в белое вещество головного мозга.
Клетки различных слоев коры объединены в «модули» – структурно-функциональные единицы. Это группы нейронов из 10–1000 клеток, которые выполняют определенные функции – «обрабатывают» тот или иной вид информации.
Каждый из слоев коры мозга значим для темпа и качества ее созревания. Они не только связаны между собой и тем самым обеспечивают целостность протекающих в коре психических процессов, но и осуществляют другие взаимодействия: межполушарные, таламо-кортикальные, кортико-стволовые, кортико-спинальные. Чем полноценнее слои коры мозга, тем больше вероятность нормативного созревания человека в эмбриональный и последующие периоды жизни.
Функциональная специализация коры мозга и его долей
Общие положенияДолгое время ряд ученых, среди них Карл Лешли, Джон Уотсон и др., считали, что кора мозга однородна, то есть каждая из ее частей в равной степени функционирует. Иными словами, различные отделы коры мозга признавались функционально эквипотенциальными, то есть обладающими одинаковым функциональным потенциалом. Следовательно, существовало убеждение, что они взаимозаменяемы. Сегодня многие взгляды на специализацию мозговой коры ушли в прошлое. Единодушно признается, что отделы мозга врожденно «запрограммированы» на разные виды деятельности, то есть они являются модально-специфичными, иными словами – функционирующими на разных анализаторных основах (слуховых, зрительных, тактильных и др.). Существует даже генетическая модальностная (анализаторная) запрограммированность нейронных сетей. Более того, наш мозг, еще ничему не обученный, владеет каким-то тайным «знанием». По меткому замечанию известного американского нейробиолога Майкла Газзаниги, человеку легко научиться бояться змей, но трудно научиться испытывать страх при виде цветов. Нобелевский лауреат Роджер Сперри также утверждал, что рост нейронных сетей является результатом генетического кодирования нейронных контактов или прочно освоенных ими программ. Сперри провел множество экспериментов, чтобы доказать это. Согласно постулируемой им модели, отростки нейронов «смотрят», куда им расти (прощупывают почву), ориентируясь благодаря вырабатываемому ими химическому веществу. Для этого нейроны как бы выпускают маленькие тонкие «щупальца» и растут в заданном от природы направлении. Обучение не в состоянии изменить или повлиять на этот процесс. Такое мнение разделяется и развивается также рядом отечественных современных ученых (К. В. Анохин). Однако благодаря множеству других исследований выяснилось, что мозг человека обладает не только свойствами, заданными генетически, но и в той же, если не в большей мере, зависит от научения – освоения окружающего мира.
Конец ознакомительного фрагмента.
Текст предоставлен ООО «ЛитРес».
Прочитайте эту книгу целиком, купив полную легальную версию на ЛитРес.
Безопасно оплатить книгу можно банковской картой Visa, MasterCard, Maestro, со счета мобильного телефона, с платежного терминала, в салоне МТС или Связной, через PayPal, WebMoney, Яндекс.Деньги, QIWI Кошелек, бонусными картами или другим удобным Вам способом.
Вы ознакомились с фрагментом книги.
Для бесплатного чтения открыта только часть текста.
Приобретайте полный текст книги у нашего партнера:
Всего 10 форматов