
Палеонтология антрополога. Три эры под одной обложкой
Оледенение было не единым, а разделяется как минимум на три отдельных этапа: рэпитенский гляциопериод – 723–709 млн л. н., стертовский – 667–658 млн л. н., марино – около 636–635 млн л. н. (понятно, все эти даты довольно условны и периодически пересматриваются). Внутри каждого гляциопериода выделяются гляциоэпохи, каждая из которых содержит отдельные ледниковые события.
Лапландское оледенение (как, кстати, и предыдущие, и последующие) в немалой степени было вызвано живыми организмами. Фотосинтетики потребляли углекислый газ, превращали его в органические вещества и в немалых количествах откладывали в осадок. Неспроста ещё в раннем протерозое сформировались огромные залежи графита, например, в Приазовье, на юге Гренландии, на Кольском полуострове и в Карелии. Тогда же продолжали образовываться исключительно богатые залежи нефти и горючих сланцев. К концу протерозоя такое массивное изымание углерода из атмосферы не могло не сказаться. Парниковый эффект снизился – оледенение было неизбежно. Не способствовало потеплению и уменьшение вулканизма, а стало быть – ослабление эндогенной дегазации. Да и вообще планета остывала, тектонические движения были уже не те, приливные деформации подкорковых оболочек сбавили обороты, приток радиоактивного тепла тоже снизился, так как за предыдущие миллиарды лет многие элементы успели распасться. Даже астероиды на планету сыпались реже, так как беспорядочного космического мусора в Солнечной системе поубавилось – что-то уже успело попадать, что-то притянулось планетами-гигантами и сконцентрировалось в астероидных поясах.
Нам повезло. Во-первых, жизнь окончательно всё-таки не замёрзла. В глубинах океана, около геотермальных источников, в подлёдных озёрах, на вершинах гор и узкой полосе открытой воды на экваторе продолжали расти и копошиться археи, бактерии, водоросли, грибы и какие-то неведомые животные. Надо думать, каждое оледенение вызывало вымирание части видов, а стало быть, высвобождение экологических ниш. Как это всегда бывает, жёсткие условия вызывали усиление отбора и ускорение эволюции с закономерным всплеском видообразования у тех, кто вымирать не хотел. Свято место пусто не бывает и нет худа без добра – диалектику никто не отменял.
Более того, в холодной воде кислород растворяется лучше, чем в тёплой (неспроста все китобои гонялись за китами не в тропиках, а в Арктике и Антарктике, где много фитопланктона, а стало быть – и зоопланктона, а стало быть – и китов), так что там, где вода не замерзала или хотя бы иногда оттаивала, плотность жизни была очень большой.
Другое следствие холода – увеличение солёности воды, ведь пресная замерзала, вытесняя соли в оставшуюся незаледеневшую. А в более плотной воде проще плавать. Кстати, это же помогало вырасти гигантским акритархам, помянутым выше. Легче жилось и многоклеточным животным.
Неспроста именно в это время появляются первые более-менее достоверные следы губок. Правда, остатки самих губок пока не найдены, но в Омане в отложениях с древностью 635–713 или даже до 751 млн л. н. выявлены следы их метаболизма в виде 24-изопропилхолестана – вещества, специфического для обыкновенных губок Demospongiae. Показательно, кстати, что губки эти жили на мелководье, а в глубоководных отложениях признаков их присутствия нет – там покамест царили безкислородные – анаэробные – условия. Но губки и, возможно, другие животные делали своё дело – фильтровали воду от взвешенной в ней органики, чистили и делали прозрачной. Больше света попадало на большую глубину, зона кислородной жизни всё больше расширялась.
Эдиакарий
635–541 миллион лет назад: Время чудес
Первым делом, стоит прояснить терминологию конца протерозоя. Иногда он называется эдиакарским периодом, иногда – вендским. Эти два слова так часто используются вместе, что многим представляются синонимами, но это не совсем так. Эдиакарий международной шкалы начался 635 млн л. н., а венд российской – 600 млн л. н.; 35 миллионов лет разницы, при всей спорности и подвижности датировок докембрия, весьма немалый срок (в схемах разных лет цифры могут ещё значительно гулять). Дело в принципах выделения: автор понятия «венд» Б.С. Соколов в 1950–1952 и 1964 гг. геологически строго определил его чёткие границы, обозначив важнейшими признаками лапландское оледенение и следы характерных мягкотелых организмов. Впрочем, не все были готовы принять докембрийские времена: первоначально венд был признан лишь как начало кембрия и назывался валдайским комплексом; в 1975 г. венд был узаконен в стандартной глобальной шкале, а в 1990–1991 гг. – в международной шкале. В последующем представления о докембрийских оледенениях бурно развивались, добавились новые характеристики, и период стал расти, по пути в 2004 г. поменяв название в международной шкале (зарубежным исследователям бывает трудно принять приоритет российских исследователей, даже когда он очевиден, тем более когда для названия периода используется название славянского племени вендов). Таким образом, сейчас венд фактически является вторым отделом эдиакария, хотя формально это не отражено ни в международной, ни в российской геохронологических шкалах. Впрочем, иногда предлагается, прямо наоборот, эдиакарий считать частью венда, что не добавляет согласия и ясности. В довершение, существует понятие «синийского периода», выделенного А. Грэбо по китайским разрезам раньше всех – ещё в 1922 г., а потому имеющего приоритет, но и огромный минус – отсутствие чётких границ.
Как бы ни называть конец докембрия и как бы ни очерчивать его границы, это было архиважное время. Материки в очередной раз собрались в суперконтинент Паннотия; в завершение африканской гляциоэры друг за другом грянули два ледниковых периода: гляциопериод гаскье около 582 млн л. н. и – на самой границе протерозоя и кембрия – байконурское оледенение 547–533 млн л. н. На протяжении эдиакария содержание кислорода в атмосфере и океане опять начало бурно расти, это называется вторым кислородным скачком. Существенно, что в разных слоях океана установились почти современные проценты кислорода, аммиака и железа – по этим параметрам венд-эдиакарий больше похож на фанерозой, а не на остальной протерозой. Особенно важно, что кислородом насытились не только поверхностные слои, как было до этого, но и большие глубины. Что было причиной, а что следствием – вопрос, но одновременно в биосфере наконец-то начинают преобладать эукариоты.
В венде широко распространяются нормальные ленточные водоросли Vendotaenia antiqua и Kanilovia insolita, обладавшие полноценным многоклеточным талломом. Есть предположение, что каниловии уже начали осваивать сушу, по крайней мере на их талломах найдены ряды странных катушек Cochleatina canilovica, которые, возможно, использовались для разбрасывания спор в воздухе. Правда, пока вопросов тут больше, чем ответов (спор никто не нашёл, некоторые «виды» катушек не связаны с водорослями, на талломах нет следов прикрепления катушек), так что утверждать что-то невозможно.
Эдиакарская фауна – великая загадка. В настоящее время её представители найдены в самых разных местах – от Австралии (где находятся Эдиакарские горы, давшие название фауне и периоду), Аргентины и Намибии, Испании и Англии до Белого моря и Урала, Забайкалья и Ньюфаундленда. Казалось бы, находок огромное количество, про них написаны сотни статей и книг, всё уже должно быть известно. Ан нет. До сих пор суть эдиакарской фауны не раскрыта.
Эдиакарские существа – вендобионты – имеют несколько странностей.
Во-первых, необычные формы симметрии. При скользящей симметрии есть правая и левая части тела и вроде бы то ли верх-низ, то ли перед-зад, тело явно сегментировано, но сегменты смещены в шахматном порядке. Самые известные существа такого рода Charnia, Spriggina, Yorgia и Dickinsonia (некоторые считают, что три последних рода – одно и то же, только разной степени вытянутости). Pteridinium, кроме скользящей симметрии, был к тому же и трёхлопастным. Обычно каждый следующий сегмент был чуть меньше предыдущего, отчего тело Charnia, например, в итоге становилось похожим на лист, перо или наконечник копья. Другой вариант – спиральная трёхлучевая симметрия у «медузоида» Tribrachidium heraldicum – круглого существа с тремя завитушками на «макушке». Впрочем, другие – Ediacaria, Tateana (известная также как Cyclomedusa) и Mawsonites – имеют строго круглую форму и радиальную симметрию, более похожую на вариант современных кишечнополостных. «Чешуйки», расходящиеся не то кругами, не то спиралью по поверхности Mawsonites spriggi, наводили многих на мысли об иглокожих. Некоторые диски могут быть заглублённым в почву основанием «перьев»; лучшим примером являются несколько видов Charniodiscus и Paracharnia, верхняя сорокасантиметровая лопасть которых похожа на чарнию, а низ представляет собой типичного «медузоида».
Во-вторых, эдиакарские твари обладали изометрическим, то есть равномерным ростом: маленькие и большие экземпляры совершенно одинаковы в своих пропорциях и отличаются лишь размером.
В-третьих, у вендобионтов нет следов органов передвижения и самих следов движения. Даже те, что напоминают червей и имеют что-то похожее на голову, как Spriggina floundersi, были, судя по всему, неподвижными существами, а «голова» на самом деле была просто первым сегментом, больше разросшимся. Между прочим, отсутствие роющих животных было одной из причин приличной сохранности отпечатков вендобионтов: никто не копался и не баламутил дно, так что осадок мог спокойно фоссилизироваться. Конечно, всегда найдутся исключения: беломорская Yorgia, судя по следам, вполне умела ползать, а горизонтальные норки в толщах строматолитовых бактериальных плёнок всё же найдены.
В-четвёртых, эдиакарские существа обычно не имеют ртов, кишечников или чего-то похожего на пищеварительную систему. Ни разу не понятно, как бы они могли питаться. Предположения о фото- или хемосинтезе законны, но как их доказать? Кстати, небогато и с органами размножения. У Dickinsonia посреди тела проходила полость с отходящими боковыми каналами, но её назначение неизвестно. Рядом с некоторыми отпечатками дикинсоний есть пятна того же размера и формы, которые, возможно, являются следами питания. Предполагается, что дикинсонии ели подобно современному трихоплаксу, медленно наползая на еду, выделяя на неё пищеварительные ферменты и всасывая получившуюся кашицу.
Намибийские Ernietta plateauensis могли питаться осмотически, впитывая осадок, попадающий на «кожу» за счёт турбулентных завихрений воды над рифлёными краями своих кубкообразных тел, а жизнь колониями помогала им в этом.
В-пятых, на самих вендобионтах нет повреждений – погрызов, покусов, оттяпанных кусков. Создаётся впечатление, что в фауне не было хищников, рисуется картина идиллического мира пацифистов.
В-шестых, вендобионты не имели никаких твёрдых частей тела – ни панцирей, ни раковин, ни зубов. Впрочем, раз они никого не кусали и их никто не грыз, им это было не нужно.
Кем были эдиакарские вендобионты? На эту темы выдвинуто огромное количество предположений: колонии бактерий, водорослей или простейших, грибы, лишайники, медузы, черви, вымершие типы, не имеющие родства с современными. Описано огромное число типов вендобионтов, но их достоверность под большим вопросом; в систематике и родстве эдиакарских существ предстоит ещё долго разбираться.
В последнее время использование тонких химических методов позволило приоткрыть завесу тайны. Так, в круглых концентрических отпечатках Beltanelliformis (они же Nemiana), собранных кучками, оказался повышен процент гопанов – веществ, в которые преобразуются составляющие бактериальных мембран. Особенно важно, что из рядом расположенных водорослевых плёнок выделены стераны – производные мембран водорослей, что подтверждает неслучайность распределения веществ в породе. Таким образом, Beltanelliformis были колониями бактерий, росших на водорослевых плёнках типа Nostoc. А вот в отпечатках Dickinsonia нашлись следы холестерина, стало быть, это были уже настоящие животные.
Как вы там, потомки?
В 2014 году случилась сенсация: у учёных дошли руки до коллекций, собранных глубоководным тралом в 1986 году в пучинах Бассова пролива у берегов Тасмании. Трал применялся не простой, а хитрый, способный захватить парящих в толще воды животных на глубине 400 – 1000 метров. И вот почти треть века в баночках на полке в музее плескались странные существа, на которых никто не обращал внимания. А зря! Твари оказались очень странными. Они были описаны как Dendrogramma enigmatica и D. discoides. Дендрограммы выглядят как грибочки-лисички на ножке; внутри стебля идёт канал, который в шляпке расходится дихотомически ветвящимся деревом, за что, собственно, зверюшки и получили имя. Такая конфигурация чрезвычайно напоминает эдиакарских Rugoconites tenuirugosus, R. enigmaticus, Albumares brunsae и Anfesta stankovskii. Да что уж скромничать – сходство с Rugoconites вообще абсолютное. Так что дендрограммы, возможно, являются уникальными реликтами эдиакарской фауны. Позже, в 2015 году, конечно, последовало разоблачение: генетический анализ новообретённых образцов показал, что тасманийские «грибочки» относятся к сифонофорам Siphonophorae семейства Rhodaliidae – давно известной группе кишечнополостных (кроме того, два описанных вида оказались единым). Но ведь это не отменяет сходства с эдиакарскими существами! Не является ли это доказательством существования сифонофор в венде?
Странный трёхдольчатый Ventogyrus chistyakovi из венда Белого моря вполне мог быть сифонофорой; по другой версии это – первый гребневик, хотя современные гребневики делятся на чётное число долек.
Кроме вендобионтов, в эдиакарии известны и другие существа. Inaria была похожа на губку с заглублённым в осадок полым, слегка сегментированным расширением и торчащей наружу трубкой десяток сантиметров длиной; судя по внутренним септам, потенциально она может быть кишечнополостным, а может – обломанным основанием кого-то типа Charnia. Arkarua adami напоминает пятилучевого иглокожего и даже иногда включается в класс Edrioasteroidea, хотя такое определение сугубо гипотетично.
В самом конце венда появляются и организмы, имевшие твёрдые покровы, особенно много найдено их в Намибии, Испании и Якутии.
Cloudina, возможно, напоминала червей, живущих в известковых трубках, но могла быть и чем-то наподобие полипа. От них сохранились цепочки вставленных друг в друга кальцифицированных конусов, причём колонии клаудин были весьма плотными и многочисленными, образуя многокилометровые рифы. В трубочках клаудин обнаружены маленькие отверстия, первоначально интерпретированные как следы нападения хищников. Однако строго квадратная форма отверстий и химический состав скелета выдаёт истину: дырочки образовались из-за нарастания, а потом выпадения кристалликов доломита. Очень похожи на клаудин Sinotubulites, формировавшие те же рифы. Вместе с клаудинами жили и Namacalathus hermanastes, тоже обладавшие кальцитовым скелетом, но имевшие совсем другую форму – тонкая трубка, увенчанная восьмигранником с отверстиями на каждой плоскости. Protolagena были похожи на гладкую круглую вазу с горлышком (возможно, как и Inaria, это – основания вендобионтов типа Charnia), якутские Suvorovella aldanica и Mayella – плоские диски, Palaeopascichnus linearis и Orbisiana simplex – членистые ветвящиеся стебли, первоначально принятые за следы ползания.
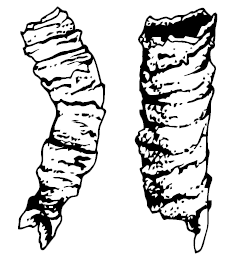
Cloudina. Эдиакарское существо с твердым покровом
Замечательно, что при ближайшем рассмотрении многие вендские существа со скелетом соответствуют таким же, но бесскелетным: Suvorovella – Aspidella и Eoporpita medusa, Cloudina – Conotubus, Sinotubulites – Corumbella, Protolagena – Sicilagena. Видимо, способность формировать домики возникала более-менее независимо и одновременно. Вопрос – зачем им это было нужно? Возможно, дело в хищниках, но, как уже говорилось, достоверных следов нападения на эдиакарских существ нет. А возможно, прочным существам было проще противостоять прибою, ведь они жили на мелководье.
Наконец, в венде появились первые билатеральные животные. Сейчас таких найдено довольно много. Вероятно, древнейшим примером является Vernanimalcula guizhouena из формации Доушаньто в Китае с древностью 580–600 млн л. н. Эти крошечные создания размером в десятую долю миллиметра уже, возможно, были не только двустороннесимметричны, но ещё и трёхслойны, обладали ртом, сквозной пищеварительной системой со ртом и анальным отверстием, а также замкнутым целомом, то есть полостью тела между двумя слоями мезодермы. В принципе, это краткое описание довольно продвинутого существа уровня кольчатого червя или моллюска, даже более крутого, чем плоский или круглый червь – вполне достаточно, чтобы считать Vernanimalcula Великим Предком.
Маленькая тонкость
Сквозная пищеварительная система – огромнейшее достижение. У кишечнополостных есть одно отверстие, ведущее в пищеварительную полость; фактически, гидра, коралловый полип или медуза – это наша гаструла, эмбриональная стадия в виде вмятого самого в себя мешочка. Такое существо может питаться только очень медленно, ведь глотать и гадить приходится через одно и то же отверстие, по очереди. К тому же при таком раскладе невозможно подразделить пищеварительную систему на отделы, во всём объёме должна быть одна и та же кислотность, по всей площади выделяются одни и те же ферменты, так что переваривать можно только какой-то один набор питательных веществ, а отходов получается очень уж много. Короче, такое питание неэффективно, обмен веществ низок, меню бедное, вкус неразвит, а жизнь скучна. Куда веселее становится, когда ротовое и анальное отверстия разделяются: теперь можно лопать без остановки, а пищеварительный тракт разделить на отрезки с разным назначением – тут мы перевариваем белки в кислой среде (назовём это желудком), а тут – жиры и углеводы в щелочной (пусть это будет кишечник). Потребление возрастает, обмен веществ растёт, жизнь обогащается новыми смыслами – появляется повод ещё кого-нибудь догнать и попробовать на вкус.
Некоторые шарообразные «акритархи» из Доушаньто – Tianzhushania spinosa, Appendisphaera tenuis и Ericiasphaera magna – оказались покоящимися яйцами многоклеточных беспозвоночных, окружёнными оболочками, а другие – Megasphaera inornata, M. ornata и Parapandorina raphospissa – эмбрионами на разных стадиях развития, причём датировки их из разных слоёв охватывают фактически весь эдиакарий. Парадокс в том, что сохранность яиц и эмбрионов фантастическая – отдельные клетки видны лучше, чем на многих современных препаратах, – а взрослых особей этих существ мы не знаем.
Беломорские Armillifera parva и Solza margarita с древностью 555 млн л. н. на первый взгляд – овальные «медузоиды» с невнятными радиальными бороздками, но при ближайшем рассмотрении в их строении выявляется право-левосторонняя симметрия и разница переднего и заднего концов, а потому они, возможно, были подвижными, хотя прямых доказательств этого пока не найдено. Эти существа могли быть моллюсками с мягкой раковиной сверху, типа современного слизня. Очень похожи, но обычно намного более вытянуты отпечатки Kimberella quadrata, найденные тоже на Белом море и в Австралии: перед передним концом кимберелл вроде бы видны следы смятого осадка, а иногда отпечатки расходятся веером, когда зверюшка ворочала «головой» в поисках более вкусных водорослей. Точно так же делают современные улитки. Вероятно, кимберелла уже имела радулу – ротовой аппарат с зубчиками для соскребания водорослей, характерные поскребы описаны под названиями Radulichnus inopinatus и Kimberichnus teruzzii. Все эти животные имели размер от пары миллиметров до пятнадцати сантиметров.
Совсем уж явно двустороннесимметричны два вида рода Parvancorina (P. minchami и P. sagitta) – разной степени вытянутости треугольные существа с явным утолщением в виде трезубца внутри или поверх тела. Парванкорина с самого начала рассматривалась как, возможно, древнейшее членистоногое и запросто может быть прямым предком трилобитов. Правда, парванкорина, в отличие от вендобионтов и примитивнейших кембрийских членистоногих, обладала аллометрическим ростом, то есть меняла форму с возрастом, но, с другой стороны, продвинутые членистоногие как раз характеризуются этой особенностью. Другие исследователи сравнивают парванкорину с видом Temnoxa molliuscula и считают чем-то вроде моллюска.
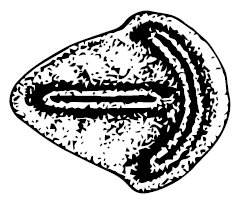
Parvancorina. Эдиакарское существо с билатеральной симметрией
Следы ползания каких-то сравнительно крупных существ по дну в виде двойных бороздок найдены в формации Такуари в Уругвае (585 млн л. н. или больше) и в долине Янцзы в Китае (541–551 млн л. н.). Уругвайские следы сплошные, там полз кто-то типа моллюска, а китайские выглядят как два плотных ряда точек, так что у оставившего их зверя, вероятно, были острые ножки. Правда, некоторые критически настроенные исследователи считают, что подобные отпечатки могли остаться и после гигантского плазмодия, но, раз в начале кембрия уже существовали вполне развитые членистоногие, то какие-то предки в конце эдиакария уже должны были водиться. Найти их более внятные останки – дело времени и упорства, а возможно, они уже и найдены. Немецкий Xenusion auerswaldae – два параллельных ряда следов – достаточно уверенно классифицируется как ксенузия, ранний вариант онихофор, довольно продвинутых созданий с парными «ножками». Вероятно, именно подобные животные оставляли парные цепочки следов в Уругвае.
Наконец, китайская Yilingia spiciformis была совсем уж откровенно билатеральной и подвижной «многоножкой», чьи многочисленные повторяющиеся сегменты были трёхдольчатыми – из центральной и двух боковых частей, а за вытянутыми телами тянулись следы ползания.
На самой границе венда и кембрия появляются Sabellidites и несколько родственных родов, которые многими исследователями уверенно относятся к кольчатым червям погонофорам Sabellida, тем более что из остатков сабеллидит выделен специфический для погонофор вариант хитина.
Маленькая тонкость
Возникновение двусторонней симметрии видится разным исследователям сильно по-разному. В целом, гипотезы разделяются на три группы.
Согласно планулоидно-турбеллярным (в том числе теория гастреи), первые многоклеточные напоминали планул – личинок кишечнополостных, то есть были двуслойными шариками с вдавленной внутрь полостью (фактически гаструлой) и одним ротовым (по совместительству и анальным) отверстием на нижней стороне. Такие живые пузырьки оседали на дно боковой стороной или ртом, начинали ползать, растягиваться и превращались в подобие современных плоских червей – турбеллярий-планарий.
По архицеломатной версии предки были похожи на четырёхлучевых кораллов, кишечные карманы которых обособились, став целомом – полостью тела, тело вытянулось, утеряв радиальную симметрию, а первичное ротовое отверстие сначала растянулось в узкую щель, а после разделилось на ротовое и анальное.
Метамерные версии (они же цикломерные) предполагают, что пращурами были многолучевые полипы, тоже начавшие ползать и вытянувшиеся, но с той разницей, что кишечные камеры дали начало сегментам-метамерам, а изначально радиальные щупальца – боковым недоножкам-параподиям. Чисто гипотетически можно представить и такой вариант, что двусторонняя симметрия возникла из двухлучевой радиальной, но это маловероятно.
Вытягивание ротового отверстия отлично видно на разных возрастных стадиях Dickinsonia, причём и по типу симметрии она представляет что-то среднее между радиальной (у молодых), скользящей и билатеральной (у взрослых). Здорово, что и сравнительная геномика показывает то же самое: у полипов все Hox-гены работают сообща вокруг ротового отверстия, а у двусторонне-симметричных животных разделяются на передние и задние. Немалое значение тут имеет решение вопроса, что у первых многоклеточных было раньше, а что появилось позже – планктонный организм, облик которого сохранился в планктонной личинке, к которой добавилась жизненная фаза донного взрослого организма (теория конечной надставки), или же донная взрослая форма, лишь потом обрётшая планктонную личинку (теория интеркаляции). Совокупность данных, учитывающих ещё и особенности формирования нервных центров (наличие его у свободно-плавающих личинок кишечнополостных и редукция у сидячих, сохранение и развитие его в мозговые ганглии у билатеральных животных), скорее свидетельствует о том, что изначальный предок был планктонным, а не прикреплённым организмом.