
Палеонтология антрополога. Три эры под одной обложкой
Спиральные разнообразнее, они включают группу типов Platyzoa, или Acoelomates: плоских червей Platyhelminthes, а также родственные типы брюхоресничных червей Gastrotricha, скребней Acanthocephala, гнатостимулид Gnathostomulida, микрочелюстных Micrognathozoa и коловраток Rotifera; группу типов Lophophorata: циклиофор Cycliophora, форонид Phoronida, брахиопод Brachiopoda, а также родственных мшанок Bryozoa и внутрипорошицевых Entoprocta (или Kamptozoa); группу типов Trochozoa: кольчатых червей Annelida, моллюсков Mollusca, немертин Nemertea и сипункулид Sipuncula.
К линяющим относятся круглые черви Nematoda, волосатики Nematomorpha, онихофоры Onychophora (известные также как лобоподы Lobopoda), тихоходки Tardigrada, членистоногие Arthropoda и ещё ряд малоизвестных типов – лорициферы Loricifera, киноринхи Kinorhyncha и приапулиды Priapula (или Priapulida).
Трудно сказать, кто из всех этих существ примитивнее. Например, плоские черви не имеют сквозной пищеварительной системы, чем похожи на кишечнополостных, зато у них два слоя мускулатуры, тогда как у круглых червей со сквозной пищеварительной системой – только продольный слой мышц. Всегда наготове мнение, что «примитивное» состояние на самом деле вторично, а предки-то были ого-го! – не то, что нынешнее племя, куда более продвинутыми. Нет определённости и в том, что было раньше – сегментированное тело или монолитное. Как бы логично, что сегментация появилась не сразу, но, во-первых, она могла возникать несколько раз параллельно (например, для кольчатых червей и членистоногих это более чем вероятно), а во-вторых, у некоторых она со временем опять пропадала, так что по крайней мере у части животных нерасчлененность вторична. Аналогично много раз параллельно возникали и менялись челюстные аппараты, кровеносная, дыхательная и выделительная системы.
Маленькая тонкость
Одно из важнейших свойств билатеральных животных – трёхслойность. Кроме наружной эктодермы и внутренней энтодермы, у них появился промежуточный слой мезодермы, то есть мышц. Специальный двигательный слой позволил билатеральным существам стать бодрыми и активными, быстро и целенаправленно перемещаться в нужном направлении, а ганглиозная нервная система со сгущениями нервной ткани обеспечила эту механику управлением.
Одно из величайших достижений кембрийских животных – глаза. Конечно, некие фоторецепторы были уже у докембрийских многоклеточных; в конце концов, светочувствительные пигменты есть даже у одноклеточных жгутиконосцев, но полноценные глаза появились только в кембрии. Ксенузии, онихофоры, тихоходки и первые хордовые имели и имеют лишь отдельные пигментные пятнышки и простейшие глазки, но среди моллюсков, динокаридид и уж тем более членистоногих глаза быстро эволюционировали, причём разными путями. У хордовых и моллюсков они сохранили простоту, но в итоге стали очень большими и обзавелись внутренними линзами. У многих членистоногих многочисленные маленькие глазки стали собираться в кучки, так что уже у аномалокариса фасеточные глаза имели диаметр до 3 см и состояли из 16 тысяч линзок. У трилобитов поверх глаз были надеты «хрустальные очки» – линзы из кальцита с магнием. Они одновременно защищали живой глаз и давали большую глубину резкости, а благодаря невероятной прочности и инертности сохранились до наших дней в изначальном виде. На многих окаменелостях стеклянные глаза трилобитов продолжают загадочно поблёскивать и переливаться желтоватым и бирюзовым цветами. Впрочем, есть сомнения, были ли «очки» прижизненными или возникали позже, в процессе фоссилизации.
Изобретение глаз в кембрии происходило параллельно много раз, так как зрение даёт огромные преимущества: оно позволяет оценить расстояние, направление движения, скорость, размер и массу прочих характеристик угрожающего хищника или, напротив, желанной добычи. Хемо- и механорецепция проигрышны, ведь на расстоянии такие органы чувств работают плохо: неизвестно, откуда принесло химию и как долго она тут, а определять опасного хищника на ощупь тем более чревато. Представьте ползущего во мраке первобытного червяка, уткнувшегося во что-то злое и поглаживающего найденное своими тентаклями: «О-о-о, какой большой и зубастый, опасный, наверное… Ой-ой…» Другое дело – глаза. Кто обладает зрением, становится королём моря! Слепые, если только не чрезвычайно многочисленны или не защищены панцирями и ядами, тут же проигрывают эволюционную гонку – либо становятся обедом зрячих, либо гибнут с голоду, не в силах поймать зоркую еду. Стоило появиться глазам у кого-то одного, как у прочих не оставалось выбора – либо тоже совершенствоваться, либо вымирать.
Из спиральных, видимо, уже в раннем кембрии существовали моллюски Mollusca. По крайней мере этим временем датируется формация Махто в канадской Альберте, в слоях которой найдены целые радулы – зубные щётки – неких неименованных моллюсков. Да и вендская кимберелла, как уже говорилось, очень уж похожа на слизняка. В немакит-далдынском ярусе Якутии найдены проблематичные микроскопические ракушечки – Purella antiqua, Oelandiella korobkovi и многие другие, подобные же окаменелости известны и из Китая. Их родство находится под большим вопросом, так как строение тела этих существ неизвестно, но не исключено, что это уже были моллюски. В том же немакит-далдынском веке уже жил моллюск Helcionella, до позднего кембрия существовало много его видов по всему миру. Интересно, что гельционелла имела одну раковину на спине, но похожую на раковину двустворок. Подобные существа иногда прямо классифицируются как моноплакофоры Monoplacophora; предположительно они были предками брюхоногих. К моноплакофорам же относится один из самых многочисленных видов в Бёрджесе – Scenella amii с приплюснутой конической раковинкой. В позднем кембрии такие недоулитки представлены уже множеством видов Kirengella и Hypseloconus.
Другие конические раковинки с поперечными валиками из томмотского века раннего кембрия Якутии и Австралии – Tannuella elata, T. elinorae, а также начала позднего кембрия Антарктиды – Knightoconus antarcticus – рассматриваются как возможные предки головоногих моллюсков.
Отлично сохранившиеся отпечатки Petalilium latus и Vetustovermis planus из Хайкоу, а также Nectocaris pterys из Бёрджеса похожи на головоногих моллюсков, общий план их строения напоминает каракатицу. Это были активные пловцы – плоские, с плавниковыми складками по бокам и двумя щупальцами спереди. В сравнении со «стандартными» моллюсками их голова слишком выделена тонкой «шеей», они чересчур пучеглазы, вдоль тела идут слишком многочисленные параллельные жабры, так что тело и плавниковые складки почти сегментированы. По совокупности черт Nectocarididae могут быть роднёй не только моллюсков, но и плоских червей, немертин и полихет.
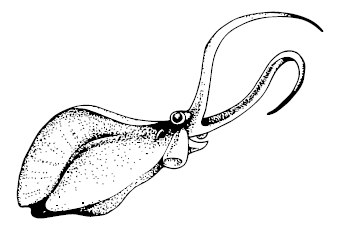
Nectocaris pterys. Моллюск
Присутствие раковины у одних потенциальных предков и отсутствие у других ставят вопрос о том, была ли она вообще у предков головоногих или же с самого начала отсутствовала, а появилась заметно позже (а потом у большинства опять исчезла)? По крайней мере, у нескольких видов позднекембрийского Plectronoceras из Техаса двухсантиметровая раковина точно была, а это существо уверенно классифицируется как первый представитель Nautiloidea. Был ли он больше похож на улитку или наутилуса, только с рогоподобной раковинкой – вопрос, но это и не так важно. Главное, с плектроноцераса началась внятная история головоногих, между прочим, мощно повлиявшая и на наше происхождение.
В канадских сланцах Бёрджес сохранились отпечатки Odontogriphus omalus – овального плоского создания, обладавшего радулой с двумя, тремя или даже четырьмя рядами зубов, которые больше всего напоминают моллюскочьи (моллюскиные? моллюсячьи? Велик и могуч русский язык, а улиток обделил!). Одонтогрифус достигал дюжины сантиметров в длину (то есть был дюжим), на спине имел мягкую недораковину, а на животе – рот; жабры-ктенидии и мантийная полость тоже выдают в нём моллюска. Зверь ползал по цианобактериальным матам Morania и скрёб их своими зубищами. Предполагалось, что это был общий предок моллюсков, кольчатых червей и брахиопод, но тщательные исследования показали наибольшее сходство этих существ именно с моллюсками. Впрочем, одонтогрифус не может быть прямым предком моллюсков и червей, так как обе эти группы появились раньше.
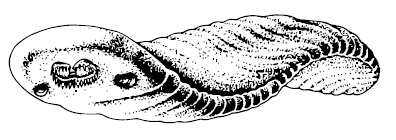
Odontogriphus omalus
Как ни странно, ближайшие родственники моллюсков – кольчатые черви Annelida. Долгое время зоологи считали их лучшими кандидатами на роль предков членистоногих, с одной стороны, и хордовых – с другой, ведь у них есть и сегменты, и щетинки, очень похожие на зачатки членистых ног, и замкнутая кровеносная система. Но реальность, как всегда, оказалась хитрее. Разделение тела на сегменты возникло, видимо, независимо у линяющих и спиральных беспозвоночных, конечности и кровеносная система тоже эволюционировали параллельно.

Canadia spinosa
Древнейшие известные кольчатые черви – Phragmochaeta canicularis из Сириус Пассет. В Бёрджесе многощетинковые черви довольно разнообразны и очень похожи на современных: самая известная зверюшка – Canadia spinosa, наиболее обычная – Burgessochaeta setigera, а самая редкостная – Insolicorypha psygma. Эволюция щетинок кольчатых червей прекрасно видна на странном существе Wiwaxia corrugata (между прочим, их отпечатки известны ещё и из Китая, и из Якутии). Эта округлая приплюснутая тварь имела плотный чешуйчатый покров, увенчанный двумя рядами высоких «зубьев» или «перьев». Выглядит это настолько странно, что долгое время виваксия не находила своего места в систематике. Но подробное исследование показало, что щетинки виваксии по сути – просто сильно увеличенные щетинки многощетинковых червей. Кроме прочего, виваксия имела радулу с тремя рядами зубов, очень похожую на радулу одонтогрифуса, несмотря на внешние отличия этих существ и вроде бы их принадлежность к разным филогенетическим ветвям.

Wiwaxia corrugata
Отношения кимбереллы, одонтогрифуса, виваксии и халькиерии (о ней речь дальше) являются предметом дискуссии. По одной версии, линия кольчатых червей отделилась от моллюсков ещё до кимбереллы, а вся указанная цепочка выстраивается в более-менее последовательный ряд: кимберелла похожа на одонтогрифуса формой тела, одонтогрифус на виваксию радулой, виваксия на халькиерию щетинками, а там и до моллюсков недалеко. По другой версии, только одонтогрифус имеет отношение к моллюскам, виваксия родственна кольчатым червям, а халькиерия – брахиоподам, а потому все они относятся совсем к разным группам беспозвоночных. Сходство же по отдельным чертам отражает более древнее родство или возникло независимо в силу конвергенции.

Haplophrentis. Хиолит
Маленькая тонкость
Конвергенция – сходство неродственных организмов, возникшее в силу похожих условий существования. Это явление чрезвычайно широко распространено в живой природе. Моллюски – брахиоподы, акулы – ихтиозавры – дельфины, черепахи – панцирные динозавры – броненосцы, а также миллионы других случаев – всё это примеры конвергенции. Подобные структуры при конвергенции могут возникать из похожих или совсем из разных зачатков.
Параллелизм – сходство родственных организмов, возникшее в силу общего происхождения. Это намного более скучное явление. Например, у всех копытных есть копыта, так как они были у общего предка; рога разных оленей при всей их вариабельности тоже суть производное рогов единого предка.
Близкая пара понятий – гомология и аналогия.
Гомологичные органы – имеющие общее эволюционное и эмбриологическое происхождение, но в итоге могущие быть похожими, а могущие – очень непохожими как по строению, так и по функции. Хрестоматийные примеры гомологичных органов – передние конечности позвоночных: рука человека и шимпанзе, передняя нога коровы, передняя лапа черепахи или крота, крыло птицы, крыло летучей мыши, грудной плавник осетра или плавник дельфина.
Аналогичные органы – схожие по внешнему виду и назначению, но возникшие независимо эволюционно и эмбриологически. Классические примеры – крылья бабочки и птицы, ноги таракана и лошади, спинной плавник тунца и кита.
Понятно, что бывают и сложные случаи. Например, головные концы мухи и мыши гомологичны, так как задаются одинаковыми Hox-генами, однако сами головы и конкретные структуры мушиных и мышиных голов аналогичны, так как возникли намного позже разделения линий членистоногих и хордовых. Общий предок имел головной конец, но не имел головы (об этом ещё пойдёт речь дальше). Проблема в том, что мы не всегда знаем, как выглядел общий предок, и уж подавно – какими генами наследуются те или иные структуры.
Ситуация усложняется существованием ещё и бёрджесских Orthrozanclus reburrus и O. elongata, выделенных вместе с несколькими родственниками в особое семейство Halwaxiidae. Ортрозанклусы сочетают черты моллюсков, кольчатых червей, брахиопод, виваксии и халькиерии. У ортрозанклуса имелась раковинка на голове, как у халькиерии, бока умеренно вытянутого слизнеподобного тела были укреплены полыми пластинками, а по краям топорщились длинные, тонкие, изогнутые и тоже полые щетинки, похожие на «перья» виваксии. Возможно, подобные ортрозанклусу беспозвоночные являлись общими предками всех указанных животных – и кольчатых червей, и моллюсков, и брахиопод. По другой же версии, учитывающей сравнительно позднее время существования, это были специализированные твари, не имевшие прямого отношения к современным группам.
Как вы там, потомки?
Внешне очень похожи на двустворчатых моллюсков брахиоподы. Вроде бы тоже две раковинки, а внутри – вытянутое тельце. Однако разница этих существ колоссальна. У моллюсков раковинки правая и левая, а у брахиопод – верхняя и нижняя. «Нога» моллюсков и «рука» брахиопод тоже устроены совершенно по-разному. Главная фишка брахиопод – фильтровальный аппарат лофофор мудрёного строения. Любопытно, что беззамковые брахиоподы сохранили сквозную пищеварительную систему, а у замковых она вторично замкнулась. Личинки имеют глазки и статоцисты, а у взрослых они исчезают за ненадобностью, что представляет отличный пример деградации. Личинки плавают очень недолго и оседают обычно рядом с родителями. Благодаря этому брахиоподы в разных местах океана сильно отличаются друг от друга, быстро специализируются и быстро изменяются, а это, в свою очередь, делает их отличными руководящими ископаемыми.
Форониды – хотя и маленькая, но крайне любопытная группа. Внешне они выглядят как перистые полипы на длинной ножке, запакованной в органическую трубочку, облепленную песчинками, но в реальности гораздо круче. У них есть вторичная полость тела; пищеварительная система сквозная, а кровеносная система замкнутая; нервная система при этом сетчатая.
Внутрипорошицевые, или сгибающиеся, внешне похожи на форонид – тоже чашечки с щупальцами на ножке, только у них анальное отверстие открывается внутри круга щупалец, нет настоящей полости тела и кровеносной системы, зато есть нервный узел; отличается и выделительная система.
Когда появились брахиоподы и их родственники, не вполне понятно. Возможно, их предки кроются в «мелкораковинной фауне» самого начала кембрия или даже конца протерозоя. В раннем кембрии известен целый ряд потенциальных предков: в китайском Чэнцзяне – странные существа Cotyledion tylodes, Yuganotheca elegans и Phlogites longus, в раннем кембрии Южной Австралии – Eccentrotheca helenia и Micrina etheridgei, в Сибири, Марокко и Гренландии – ряд видов Tannuolina. Родство их спорно, но есть мнение, что котиледион был внутрипорошицевым, эццентротека – форонидой, юганотека – не то форонидой, не то переходным звеном от хиолитов к брахиоподам, флогитес – представителем особого типа Dendrobrachia, сочетающего черты внутрипорошицевых, форонид и мшанок (Cheungkongella ancestralis описана как древнейшая асцидия, но при ближайшем рассмотрении оказалась тем же флогитесом), а микрина – вполне себе предком брахиопод. Внешне они были довольно разнообразны: котиледон выглядел как кубок на длинной ножке, покрытый множеством крошечных раковинок, эццентротека – тоже как кубок, но без ножки и облепленный более крупными раковинками, флогитес – как кубок на короткой ноже, но без раковинок, зато с щупальцами, юганотека – как длинный червяк, приделанный к рожку мороженого, который вместо шариков пломбира прикрывали две овальные раковины, а таннуолина – как короткий широкий червь с двумя плосколежащими на спине ракушками, правой и левой. Намного больше на современных брахиопод были похожи раннекембрийские Heliomedusa orienta из Китая и близкая Mickwitzia occidens из Северной Америки, а Lingulosacculus nuda из Альберты сочетает черты брахиопод и хиолитов. Dinomischus isolatus и D. venustus из Чэнцзяна, а также Siphusauctum lloydguntheri и S. gregarium из начала среднего кембрия Юты и Бёрджеса – придонные «тюльпаны» на тонкой двадцатисантиметровой ножке. К кому ближе эти фильтраторы – совершенно непонятно, но есть мнение, что они родственны внутрипорошицевым.
Другой потенциальный вариант возникновения брахиопод вырисовывается из строения Halkieria evangelista из Сириус Пассет в северной Гренландии. Халькиерия регулярно упоминается как предок кольчатых червей, моллюсков и брахиопод, но реальное отношение имеет, видимо, только к последним. Она выглядела как шестисантиметровый вытянутый слизняк с двумя раковинками на спине – над передним и задним концами. Предполагают, что такой зверь постепенно укорачивался, потом сложился пополам, и так получилась брахиопода с верхней и нижней раковинами. Впрочем, строение и расположение щетинок, покрывающих тело халькиерии, чрезвычайно напоминают таковые у виваксии, только масштаб совсем другой, так что линии кольчатых червей и брахиопод в среднем кембрии разошлись ещё не так сильно; другие же исследователи видят в халькиерии и потенциального предка моллюсков, хотя хронологически моллюски появились раньше.
В среднем кембрии есть уже настоящие брахиоподы; например, в Бёрджес это Acrothyra gregaria, Linguella waptaensis, Micromitra burgessensis и многие другие. Некоторые раковины просверлены каким-то хищником, что наглядно показывает, во-первых, причину возникновения раковины, а во-вторых, бесполезность извечной борьбы средств защиты и нападения. В конце кембрия брахиопод уже было величайшее изобилие.
В позднекембрийских слоях Мексики найдена и древнейшая мшанка – Pywackia baileyi, но расцвет этой группы случился уже в следующем периоде.
Хиолиты Hyolitha – загадочные родственники брахиопод, защищённые острой конической раковиной с гранёными краями и закрывавшиеся крышечкой, из-под которой изгибались два тонких «бивня». Древнейшие хиолиты Paragloborilus subglobosus известны из раннего кембрия Якутии; в Бёрджесе найдены Haplophrentis carinatus и H. reesi, бывшие добычей для Ottoia. Долгое время хиолиты считались моллюсками или ближайшими их родственниками, но открытие у них лофофора со щупальцами расставило всё на свои места.
* * *Линяющие представлены меньшим числом типов, но большим – родов и видов.
Примитивнее всех выглядят круглые черви и волосатики, но, к сожалению, они почти не известны в ископаемом виде. Возможно, Maotianshania cylindrica и Markuelia – древнейшие представители волосатиков Nematomorpha, но пока это только предположение.
Как вы там, потомки?
Современные волосатики – паразиты членистоногих, чаще всего насекомых, иногда ракообразных, причём меняют хозяев. Видимо, изобилие членистоногих в кембрии уже вызвало появление и специфических паразитов. Интересно, что взрослые волосатики мало отличаются от круглых червей и внешне выглядят как длинный тонкий двигающийся волос, тогда как личинки очень похожи на приапулид – такие же толстые получленистые черви с выдвигающимся когтистым хоботком. Таким образом, волосатики могут быть куда более продвинутой группой, взрослые формы которых упрощены вторично.
Следующий уровень развития берут Xenusia; иногда ксенузии рассматриваются как предки онихофор и приапулид, иногда как подгруппа онихофор или, в более широком смысле, лобопод, иногда как искусственная сборная группа. Эти существа зависли между нерасчленённостью древнейших многоклеточных и сегментированностью членистоногих. План строения ксенузий всегда схож: по бокам червеобразного, слабо подразделённого тела отходили пары длинных толстых сосископодобных «ножек», которые благодаря почтисегментированности могли втягиваться или, наоборот, вытягиваться (альтернативное название «Lobopoda» буквально переводится как «дольконогие»). Особенно хорошо узнаваемы крючки на кончиках лапок. Некоторые такие коготки, например, Mongolodus, долгое время числились конодонтами. Разные представители отличаются количеством и длиной «ног», сложностью развития головы, например, строением хоботка, наличием или отсутствием щупалец и глаз. Очевидно, в кембрии они переживали расцвет, не то что их нынешние потомки онихофоры.
Одни из древнейших ксенузий из Чэнцзяна – двухсантиметровые Microdictyon sinicum, одни из самых примитивных, хотя и не самые ранние – Ovatiovermis cribratus из Бёрджеса. Известно и огромное количество других родственных существ из самых разных мест планеты – Luolishania, Cardiodictyon, Mureropodia, Paucipodia, Antennacanthopodia, глазастая Miraluolishania haikouensis и прочие.
Самые известные ксенузии, без сомнения, – Aysheaia pedunculata и Hallucigenia sparsa (есть и другие, более древние виды — H. hongmeia и H. fortis из Китая, но они не столь колоритны). Айшеайя, несмотря на позднюю датировку, наиболее примитивна и одновременно максимально похожа на современных онихофор. Вероятно, от подобных ей существ и возникли все остальные ксенузии, онихофоры, тихоходки и членистоногие.
Название галлюцигении говорит само за себя. Её вытянутое пятисантиметровое червеобразное тело с одной стороны заканчивалось довольно тяжёлой головой с вроде бы глазами, из тонкой шеи в стороны тянулись три пары щупалец, на животе было семь пар толстых недоножек с фирменными коготками, а на спине – тоже семь пар очень длинных шипов. Эти пики при большом увеличении чрезвычайно красивы, так как покрыты мельчайшим чешуйчатым узором; между прочим, такие щетинки давно были известны как конодонты Rhombocorniculum. Примечательно, что от ранних китайских видов к позднему канадскому длина шипов росла, стало быть – была надобность, хищники тоже хотели кушать.
Работа над ошибками
В первом исследовании спинные шипы были приняты за ходульные негнущиеся конечности, а настоящие «ноги» были ошибочно опознаны как непарные выросты на спине. На отпечатке-то верх и низ не указаны! Лишь разглядывание в микроскоп коготков на лапках позволило установить истину и сориентировать зверюшку как положено.
Ещё одна важная ксенузия – Diania cactiformis из китайского Хайкоу. Десять пар её недоног были покрыты множеством коротких шипов и при этом заметно толще тонкого тела, так что всё животное походило на ветку араукарии. Самое же важное, что при общем плане строения, как у всех ксенузий, и тело, и «ноги» диании были отчётливо членистыми, так что это существо лучшим образом подходит на роль общего предка всех членистоногих. Правда, подкачала «голова» диании – безглазый сегментированный баклажан. Другое дело – якутский Siberion lenaicus из Синска, имевший огромную растопыренную переднюю пару «ног», усаженную здоровенными шипами – отличную основу для «рук» керигмачелы; похож на него и более древний Megadictyon из Гренландии и Китая.
Совершенно другую специализацию имел китайский Facivermis yunnanicus, «ноги» которого превратились в пять пар перистых щупалец на переднем конце очень длинного тонкого тела, благодаря чему изначально он был принят за многощетинкового червя.
Возможно, ксенузии, перешедшие к роющему образу жизни, окончательно потеряли свои «ноги» и стали приапулидами, те, что стали активно плавать и клацать передними зубастыми ногочелюстями, превратились в аномалокарид, вышедшие на сушу и лучше сохранившие исходный план строения – в онихофор, а зависшие в капельках воды на водорослях и мхах – в тихоходок.