
Эндокринные синдромы и болезни. Руководство для врачей
В этом классическом определении термины эндокринная железа и гормон взаимозависимы. Отсюда очевидна в клинической эндокринологии логика диагностического поиска – через исследование гормонов крови диагностировать болезни эндокринных желёз.
3.1.2. Определение эндокринной железы
Эндокринная железа – чётко очерченная макроанатомическая структура, главной функцией которой является синтез биологически активных веществ, называемых гормонами. В клинической эндокринологии выделяют семь эндокринных желёз, функции которых оценивают по исследованию в крови вырабатываемых железой гормонов. Для оценки её функций используют не весь спектр гормонов железы, а строго ограниченный их набор, с помощью которого определяют функцию эндокринной железы (табл. 3.1). Кроме гормонов, для диагностики болезней можно использовать их метаболиты, которые иногда оказываются более надёжным маркёром эндокринной болезни, чем исследование самих гормонов. Так, в диагностике феохромоцитомы надёжнее исследование метаболитов катехоламинов метанефринов, чем адреналина и норадреналина.
Таблица 3.1.
Эндокринные железы и синтезируемые ими гормоны, которые используют для диагностики эндокринных болезней


Исследование гормонов для диагностики эндокринных заболеваний не всегда оправдано. Наиболее яркий пример – сахарный диабет, в диагностике которого не используют исследование инсулина, хотя заболевание вызвано инсулиновой недостаточностью. Также исследование окситоцина и вазопрессина не используют для диагностики их недостаточной или избыточной секреции, а нарушение их синтеза определяют по их метаболическим эффектам.
Более того, в диагностике эндокринных болезней могут быть использованы гормоны, которые не синтезируются эндокринными железами, например инсулиноподобный ростовой фактор 1 (ИРФ-1), который образуется в печени под действием СТГ. Его используют для диагностики акромегалии, вызванной опухолью гипофиза.
Синтез гормона эндокринной железой может быть:
• единственной её функцией (например, передняя доля гипофиза, щитовидная и паращитовидные железы, надпочечники);
• совмещён с генерированием половых клеток (например, яичники и тестикулы);
• совмещён с экзокринной секрецией (например, поджелудочная железа);
• совмещён с депонированием гормонов, синтезируемых за её пределами (например, задняя доля гипофиза, накапливающая окситоцин и вазопрессин гипоталамуса).
Эндокринная железа способна синтезировать:
• единственный гормон, что встречается редко (например, паращитовидная железа);
• спектр гормонов (как правило):
➧ специализированными клеточными субструктурами, в частности в надпочечниках две клеточные субструктуры – кора и мозговой слой – вырабатывают стероидные гормоны и катехоламины соответственно;
➧ отдельными клетками, объединёнными или нет в изолированные комплексы, например в гипофизе определённые гормоны синтезируют отдельные клетки (соматотрофы, лактотрофы, кортикотрофы, тиротрофы, гонадотрофы), которые не объединены в отчётливые клеточные образования; в поджелудочной железе инсулин и глюкагон продуцируются β- и α-клетками, объединёнными в островки Лангерганса.
3.1.3. Природа и функции гормонов
Гормоны разделяют на две основные группы.
• Полипептиды или производные аминокислот (большинство):
➧ сложные полипептиды [ЛГ, хорионический гонадотропин человека (ХГЧ)];
➧ пептиды средних размеров (инсулин и глюкагон);
➧ малые пептиды (тиреотропин-рилизинг-гормон – ТРГ);
➧ дипептиды (Т4 и Т3);
➧ производные отдельных аминокислот (катехоламины, серотонин и гистамин).
• Производные холестерина – стероиды двух типов:
➧ с интактным стероидным кольцом (стероиды надпочечников и гонад);
➧ с разобщённым кольцом В (витамин D и его метаболиты). Выделяют четыре основные функции гормонов в организме:
• размножение;
• рост и развитие;
• поддержание постоянства внутренней среды;
• производство, утилизация и сохранение энергии.
Отдельный гормон, с одной стороны, может оказывать разные биологические эффекты на различные органы, а в одном органе – в разное время; с другой стороны, некоторые биологические процессы находятся под интегральным контролем нескольких гормонов.
Гормоны регулируют функции следующих своих мишеней:
• другие эндокринные железы (например, связь гипофиз-надпочечник);
• функциональные системы (например, катехоламины и сердечно-сосудистая система);
• органы (например, Т4 и функция сердца или Т4 и функция мозга);
• ткани (например, кортизол и костная ткань).
3.1.4. Синтез, хранение и секреция гормонов
Пептидные гормоны синтезируются по тому же механизму, что и любые другие белки. Часто сначала синтезируется большая молекула прогормона, которая затем преобразуется в гормон меньших размеров. К примеру, препропаратиреоидный гормон пропаратиреоидный гормон паратиреоидный гормон. С другой стороны, стероиды и катехоламины синтезируются из меньших молекул.
Эндокринные органы не являются уникальным местом синтеза гормонов (к примеру, ХГЧ синтезируется практически во всех тканях организма), вместе с тем только в них синтез гормонов и его регулирование происходят наиболее эффективно. Три основные особенности отличают эндокринный орган от неэндокринной ткани, синтезирующей какой-либо гормон:
• скорость синтеза намного выше в эндокринном органе;
• превращение прогормона в гормон осуществляется только в эндокринном органе (например, проопиомеланокортин превращается в АКТГ в гипофизе, но не в мозге);
• эндокринные железы снабжены механизмом транспорта гормона в кровь, который, как правило, регулируем.
Скорость секреции гормона железой определяется скоростью его синтеза, которую могут регулировать другие, тропные по отношению к этой железе гормоны. За исключением Т4 и 1,25-дигидроксихолекальциферола, запасы гормонов в организме очень ограничены.
Стимуляция секреции гормона связана с деполяризацией клеточной мембраны и открытием кальциевых каналов, что приводит к поступлению кальция в клетку, где он соединяется с кальций-связывающим протеином. Повышение внутриклеточной концентрации кальция необходимо для активации экзоцитоза – процесса, в результате которого содержимое внутриклеточных везикул (гормонов, трансмиттеров) поступает в межклеточное пространство.
3.1.5. Транспорт и элиминация гормонов
Секретируемые в кровь гормоны транспортируются к тканям-мишеням.
• Стероиды и тиреоидные гормоны циркулируют в крови в связанном с белками плазмы состоянии.
• Полипептидные гормоны и катехоламины находятся в крови в свободном состоянии.
На уровне клеток гормоны взаимодействуют со специфическими рецепторами, расположенными:
• на клеточной мембране (кортикотропин, пролактин, ТТГ и ПТГ) – стимулируют образование циклического аденозинмонофосфата (цАМФ), который, в свою очередь, активирует протеинкиназу, участвующую в синтезе белка;
• в цитоплазме (кортизол, тестостерон и эстрадиол) – образуют комплекс с рецептором, который транспортируется в ядро, где связывается с хроматином; это в конечном итоге также изменяет синтез белка.
Гормоны элиминируются из крови в результате метаболических процессов, например пептидные гормоны инактивируются протеолитическими ферментами. В печени гормоны соединяются с глюкуроновой кислотой и секретируются с жёлчью, но при этом частично реабсорбируются, включаясь в так называемый энтеропечёночный цикл. Гормоны также выводятся с мочой.
Небольшие молекулы гормонов (Т4, в частности) связываются с белками крови, что замедляет их элиминацию из крови и поддерживает небольшой пул свободного гормона в крови на необходимом уровне. Связывание с белками также облегчает транспорт жирорастворимых стероидов.
3.1.6. Гормональные рецепторы
Рецепторы гормонов – клеточные белки, которые связывают гормон. Они находятся на клеточной мембране, в цитоплазме или ядре и реализуют две функции гормонов на уровне клетки:
• селективность;
• взаимодействие с эффекторными механизмами клетки.
Взаимодействие с гормоном вызывает в рецепторе конформационное изменение, что активирует специфическую клеточную ферментную систему, которая фактически и реализует характерный эффект гормона (рис. 3.1). Когда гормон соединяется с рецептором клеточной мембраны, в цитозоле появляются так называемые вторые мессенджеры (первый – гормон), которые, в свою очередь, активируют ферментную систему клетки. В ядре клетки гормон-рецепторный комплекс связан с дезоксирибонуклеиновой кислотой (ДНК) и регулирует экспрессию гена. Максимальный эффект гормона обычно проявляется уже в том случае, если связано менее 50 % рецепторов. Освободившиеся от связи с гормоном свободные рецепторы снова возвращаются в цитозоль или на мембрану клетки, где продолжают участвовать в гормон-рецепторном взаимодействии.
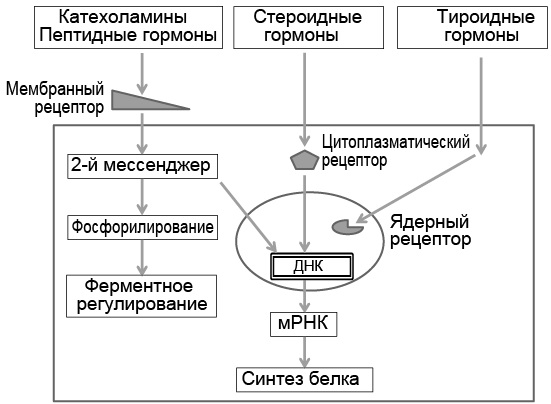
Рис. 3.1. Взаимодействие гормонов с клеточными рецепторами цитоплазматической мембраны, цитоплазмы и ядра. ДНК – дезоксирибонуклеиновая кислота; мРНК – матричная рибонуклеиновая кислота
Стероидные гормоны липофильны, поэтому свободно диффундируют через клеточную мембрану, а затем связываются с цитозольными рецепторными белками. Образовавшийся комплекс «стероидный гормон – рецептор» перемещается в ядро клетки, где он обратимо связывается с геном, изменяет ДНК, что модулирует транскрипцию матричной рибонуклеиновой кислоты (РНК).
Т3 связывается с ядерными рецепторными белками, и комплекс Т3-рецептор, соединяясь с ДНК, стимулирует образование матричной РНК. Нередко стероидные и тиреоидные гормоны действуют синергично, взаимно усиливая специфические эффекты (потенцируют экспрессию гена).
Количество рецепторов клеточной мембраны и внутриклеточных рецепторов изменяется, меняется и сила их связи с гормоном. Клетки миометрия и молочных желёз содержат окситоциновые рецепторы, число которых возрастает под действием эстрогенов (up-регулирование) и снижается под действием прогестерона (down-регулирование). В миокарде содержатся норадреналиновые рецепторы (β1), число которых и сродство к норадреналину повышается под действием тиреоидных гормонов (Т3/Т4).
Водорастворимые гормоны (моноамины, аминокислоты и пептиды) связываются с рецепторами мембраны, которая насыщена липидами, поэтому не позволяет свободно диффундировать водорастворимым гормонам через мембрану. В гормональной реакции клетки водорастворимые гормоны называют первыми мессенджерами. В ответ на их взаимодействие с рецептором внутри клетки активируются так называемые вторые мессенджеры – цАМФ, циклический гуанозинмонофосфат, инозитолтрифосфат, ионы кальция, диацилглицерол и др. Ионы кальция служат очень важным вторым мессенджером. Поток ионов кальция через клеточную мембрану в цитозоль контролируется гормон-рецепторной связью, нервными стимулами или модифицируется другими вторыми мессенджерами.
Концентрация гормонов в большинстве случаев составляет 10–10 моль/л. При этом связывание одной молекулы с мембранным рецептором приводит к образованию 10 000 молекул цАМФ в клетке, и в этом отношении цАМФ выступает в качестве молекулярного усилителя гормонального сигнала (в 10 000 раз!). Фосфодиэстераза разрушает цАМФ, поэтому её ингибиторы – теофиллин и кофеин – действуют синергично с гормонами, у которых цАМФ является вторым мессенджером. цАМФ стимулирует катаболические процессы – липолиз, гликогенолиз (глюкагон), глюконеогенез и кетогенез, секрецию инсулина β-клетками поджелудочной железы.
Глава 4
Типы саморегуляции эндокринной системы
Эндокринная система, с одной стороны, осуществляет регулирующее влияние на различные органы и ткани, а с другой – это достаточно сложная структура, которая для нормального функционирования должна обладать механизмами саморегуляции. Выделяют аутокринное, паракринное и эндокринное саморегулирование. В эндокринной саморегуляции выделяют прямую и обратную связи, причём каждая из них может быть положительной или отрицательной.
Функция эндокринной системы тесно связана с биологическими часами, т. е. она обычно циклическая. При этом биоритмическая активность эндокринной железы может быть разной – от нескольких минут до сезонных колебаний.
4.1. Аутокринное, паракринное и эндокринное саморегулирование
Молекулы гормонов относятся к большому семейству сигнальных молекул.
Аутокринные и паракринные сигнальные молекулы, вырабатываемые клеткой, секретируются в окружающую интерстициальную жидкость. Регулирующее действие этих молекул ограничивается или клеткой, которая секретировала сигнальную молекулу (аутокринное влияние), или соседними клетками (паракринное влияние).
Если эндокринная железа состоит из групп клеток, синтезирующих различные гормоны, то обычно наблюдается паракринное регулирование и между ними.
Если одна эндокринная железа (например, гипофиз) регулирует функцию другой железы (например, надпочечника), то это называется эндокринным саморегулированием.
Таким образом, термины «аутокринное», «паракринное» и «эндокринное» отражают расстояние действия молекулы гормона – от самого короткого (аутокринное) до самого длинного (эндокринное).
Пример аутокринного регулирования – секреция инсулина в зависимости от концентрации инсулина в окружающей β-клетку среде. Чем больше концентрация инсулина в окружающей среде, тем меньше секреция инсулина β-клеткой. Это свойство используют в клинической эндокринологии для диагностики инсулиномы, когда большую дозу инсулина вводят внутривенно и отслеживают секрецию инсулина на это введение. В норме секреция инсулина падает, а при инсулиноме (автономно функционирующей опухоли) она не меняется.
Пример паракринного регулирования – взаимодействие α- и β-клеток островка Лангерганса: инсулин, поступающий в интерстициальную жидкость островка из β-клетки, тормозит секрецию глюкагона α-клетками. В этом примере инсулин – паракринный регулятор.
Примером эндокринной саморегуляции в системе «гипофиз-кора надпочечников» служит стимуляция секреции кортизола надпочечниками под действием АКТГ гипофиза.
Комбинированное ауто-, пара– и эндокринное регулирование типично для гормонов: часто гормон регулирующе действует по всей длине своей системы регуляции – от короткой (аутокринной) до длинной (эндокринной).
4.2. Биоритмы
Биоритмы секреции гормонов – характерная черта функционирования эндокринной системы, и нарушение биоритма сопровождает ряд эндокринопатий. Частота ритма может составлять несколько минут или часов (пульсирующая секреция ЛГ и тестостерона), день (циркадный ритм секреции кортизола), недели (менструальный цикл) и даже более длительные периоды (сезонные изменения синтеза Т4).
4.3. Принципы прямой и обратной связи
Влияние гормона регулирующей железы на секрецию гормона регулируемой железы называют прямой связью. Влияние гормона регулируемой железы на секрецию гормона регулирующей железы – обратная связь. Если это влияние стимулирующее – связь положительная, а если подавляющее – отрицательная (рис. 4.1).
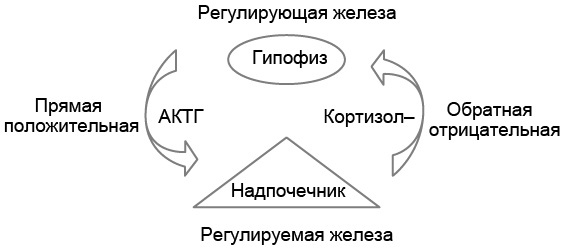
Рис. 4.1. Регулирующая железа гипофиза синтезирует адренокортикотропный гормон (АКТГ), который стимулирует образование кортизола в регулируемой железе надпочечнике (прямая положительная связь), а кортизол надпочечников подавляет секрецию АКТГ гипофиза (обратная отрицательная связь)
Отрицательная обратная связь направлена на снижение уровня гормона регулирующей железы. Так, в ответ на стресс повышается уровень АКТГ, который стимулирует секрецию кортизола надпочечниками (прямая положительная связь). В свою очередь, повышение уровня кортизола в крови подавляет секрецию АКТГ гипофизом (обратная отрицательная связь), а это, в свою очередь, ведёт к снижению концентрации кортизола в крови до предстрессового уровня. В итоге уровень управляемого гормона (в нашем примере – кортизола) всегда возвращается после значительного отклонения в некоторый заданный диапазон (базальное состояние). Почти все системы гормональной регуляции поддерживают гомеостаз гормонов путём отрицательной обратной связи. С физиологической точки зрения это очень целесообразный механизм, исключающий, с одной стороны, беспредельную стимуляцию секреции гормона регулируемой железы, а с другой – атрофию регулируемой железы.
Положительная обратная связь усиливает любую первичную стимуляцию регулируемой железы – феномен самовозбуждения системы, который встречается редко. Важный пример такой связи – повышение в крови уровня эстрадиола в середине менструального цикла. Если высокое содержание эстрадиола поддерживается более 35 ч, то активируется положительная обратная связь между яичниками и гипофизом и в результате стимулируется секреция ЛГ и ФСГ гипофиза, что, в свою очередь, приводит к дополнительной стимуляции секреции эстрадиола яичниками и т. д. В противоположность этому, более низкий уровень эстрадиола, наблюдаемый в другие моменты менструального цикла, активирует отрицательную обратную связь между яичниками и гипофизом.
Обратную связь также подразделяют на длинную (например, между надпочечниками и гипофизом), короткую (например, между гипофизом и гипоталамусом), а также авторегулирующую (секретируемый гормон влияет на синтез гормона клеткой, которая его же и производит).
Глава 5
Патофизиология эндокринной системы: клинический аспект
5.1. Причины эндокринопатий
Причины эндокринопатий могут быть разделены на пять основных категорий:
• недостаточный или избыточный синтез гормона;
• синтез патологического гормона;
• резистентность к биологическому действию гормона;
• нарушение транспорта или обмена гормона;
• множественные гормональные нарушения.
Клинические проявления этих нарушений достаточно характерны и отражают усиление или снижение физиологического действия гормона. К примеру, изменение массы тела в ту или иную сторону связано с влиянием гормонов на метаболизм – анаболизм или катаболизм. АД также регулируется гормонами: кортизолом, альдостероном и катехоламинами. Повышенная секреция этих гормонов при синдроме Иценко – Кушинга, первичном альдостеронизме или феохромоцитоме проявляется артериальной гипертензией. С другой стороны, артериальная гипотензия наблюдается у больных с гипокортизолизмом, гипоальдостеронизмом или симпатической дисфункцией. Гормоны влияют на энергетический потенциал и функцию мышц, поэтому их недостаток сопровождается такими симптомами, как быстрая утомляемость и мышечная слабость. Поскольку менструальный цикл регулируется гонадотропинами и другими гормонами, то при ряде эндокринопатий развивается менструальная дисфункция.
Указанные в предыдущем разделе физиологические модели саморегуляции эндокринной системы трансформируются в определённые патофизиологические модели, которые и служат основой для интерпретации отклонений содержания гормонов в крови от нормы. Разберем их в этом разделе.
5.2. Одноуровневая (автономная) регуляция
Если функция эндокринной железы не регулируется другой железой, т. е. она работает автономно, тогда для интерпретации результатов исследования её гормонов не используют модели саморегуляции эндокринной системы. В частности:
• пониженная концентрация гормона в крови свидетельствует о поражении эндокринной железы со сниженной секрецией гормона (например, железа удалена);
• повышенное содержание гормона в крови свидетельствует о заболевании железы с повышенной секрецией гормона (например, опухоль, секретирующая большое количество гормона).
Регуляция такого типа – одноуровневая, или автономная, поскольку она не управляется другой железой.
Если секреция гормона какой-то железы не управляется какой-либо другой железой, то это не значит, что её секреция не регулируется вовсе. К примеру, секреция инсулина поджелудочной железой не управляется другой железой, однако она зависит от концентрации глюкозы в крови, т. е. регуляция всё равно существует.
5.3. Двухуровневая регуляция
Для интерпретации результатов исследования гормонов крови двух эндокринных желёз – регулирующей и регулируемой – используют модель двухуровневой регуляции, где первый уровень – регулируемая железа, второй – регулирующая железа (рис. 5.1). Такую регуляцию осуществляет исключительно гипофиз, поэтому регулирующую железу называют центральной, а регулируемую – периферической.
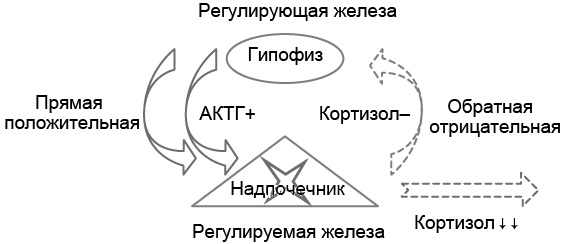
Рис. 5.1. Патофизиологическая двухуровневая модель развития гипокортицизма вследствие поражения надпочечника (периферической железы), вызвавшего гипосекрецию кортизола и повышенную секрецию адренокортикотропного гормона (АКТГ, первичное поражение периферической железы, первичная надпочечниковая недостаточность)
В этой системе регуляции главный объект исследования – функция периферической железы, так как клинические проявления эндокринной болезни являются результатом нарушения секреции её гормона, а не гормона управляющей железы. Его используют для полноценной интерпретации результата исследования гормона периферической железы, в частности топической диагностики болезни – на первом (периферическом) или втором (центральном) уровне.
Итак, исследование гормона периферической железы может дать один из двух результатов: пониженное (гипосекреция) или повышенное (гиперсекреция) его содержание в крови/моче. Рассмотрим интерпретацию этих данных с учётом двухуровневой модели регуляции.
5.3.1. Гипосекреция
В качестве примера патофизиологической двухуровневой модели рассмотрим регуляцию гипофизом функции надпочечников, которую используют для интерпретации сниженного уровня кортизола крови (см. рис. 5.1). В норме АКТГ гипофиза стимулирует секрецию кортизола надпочечниками, а повышение уровня кортизола в крови, в свою очередь, подавляет секрецию АКТГ гипофизом, т. е. прямая связь между гипофизом и надпочечниками положительная, а обратная (надпочечники гипофиз) – отрицательная.
Пример 1. У больного снижение секреции кортизола надпочечниками связано с их полным разрушением (например, при аутоиммунном процессе). Вследствие этого уровень кортизола в крови снижен. Низкое содержание кортизола в крови снимает его тормозящее влияние на секрецию АКТГ гипофизом, и концентрация АКТГ в крови значительно повышается. Эта реакция гипофиза на низкий уровень кортизола не патологическая: при временном снижении содержания кортизола высокий уровень АКТГ стимулирует надпочечники, способствуя быстрой нормализации концентрации кортизола крови. Однако в примере поражение надпочечников необратимо, потому высокий уровень АКТГ не может восстановить секрецию кортизола. Так как содержание кортизола в крови постоянно снижено, то, в соответствии с двухуровневой моделью регуляции, концентрация АКТГ будет постоянно высокой (см. рис. 5.1).
В примере поражённая периферическая (регулируемая) железа – надпочечники – не секретирует кортизол, потому его содержание в крови снижено. В ответ на сниженную концентрацию кортизола в крови секреция АКТГ гипофизом значительно повышается вследствие отрицательной обратной связи между надпочечниками и гипофизом.
Пример 2. У больного удалён гипофиз (гипофизэктомия), вследствие этого секреция АКТГ снижена и, соответственно, уровень АКТГ в крови понижен (рис. 5.2). При низкой концентрации в крови АКТГ секреция кортизола надпочечниками не стимулируется, вследствие этого содержание кортизола в крови снижено. В ответ на снижение уровня кортизола в крови секреция АКТГ гипофизом должна бы стимулироваться. Однако у больного произведена гипофизэктомия, следовательно, АКТГ не вырабатывается в принципе, и его концентрация в крови остаётся пониженной, несмотря на низкий уровень кортизола.
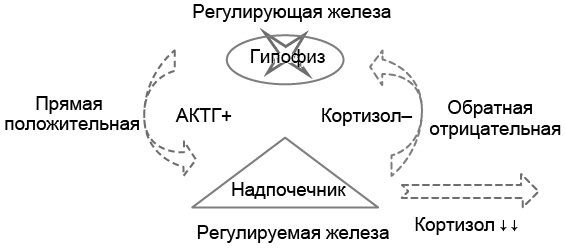
Рис. 5.2. Патофизиологическая двухуровневая модель развития гипокортицизма вследствие поражения гипофиза, вызвавшего гипосекрецию адренокортикотропного гормона (АКТГ). Снижение секреции гормона регулируемой (периферической) железы вследствие поражения регулирующей железы (вторичная гипофункция периферической железы)