
Палеонтология антрополога. Том 3. Кайнозой
Вот у кого рога развились, так это у Brontotheriidae, большая часть которых известна из Северной Америки; между прочим, филогенетически бронтотерии родственнее лошадям, а не носорогам. Самые первые мелкие Eotitanops ещё имели простенькую морду, чуть более поздние Palaeosyops размером с корову и ещё более мощные Manteoceras уже обзавелись вздутием на носу, а Protitanops curryi, Protitan имели хотя бы и невысокие, зато раздвоенные рога на носу. Бронтотерии, начав, как и все, с размера карликовой свинки, быстро выросли в огромных чудищ до двух с половиной метров в холке, став самыми большими и массивными животными своего времени. Огромные остистые отростки первых грудных позвонков были нужны для крепления мощнейшей мускулатуры для поддержания тяжеленной головы, тому же соответствовали и толстые ноги. В итоге вид бронтотериев был куда как харизматичный. Самый широко-разрекламированный бронтотерий – Megacerops (он же Brontotherium и Brontops), хотя пара его тупых округлых рогов на оригинальных черепах выглядит далеко не так впечатляюще, как на реконструкциях. У самцов рога были больше, чем у самок, что уже намекает на половой отбор и брачные турниры. Как и в случае с уинтатериями, бодание костными рогами должно было создавать сложности для сохранности мозга; впрочем, миллионы лет бронтотериям это проблемой не казалось. Известен скелет бронтотерия с результатом неудачного брачного турнира – сломанным и заросшим с костяной мозолью ребром на правом боку.

Megacerops
Монгольские Embolotherium пошли куда дальше: их носовой рог стал широченной раздвоенной лопатой, причём одинаково большой и у самцов, и у самок. Борозда на передней стороне шла от носовой полости и, возможно, служила для размещения резонатора: монгольские болота оглашались трубными кликами эмболотериев. Судя по румынскому Brachydiastematherium transylvanicum, подобные монстры жили и в Европе.
Эволюция лошадей – классика палеонтологии, каждый видел в школьном учебнике схему их эволюции, составленную В. О. Ковалевским. А начинается она с североамериканского Eohippus angustidens – первого представителя семейства Equidae. Эогиппус очень мало отличался от гиракотерия (а иногда они даже объединяются): всего 30 см в холке, со среднюю собачонку. Коронки его зубов были низкими, на ногах, как у всех примитивных непарнокопытных, по четыре пальца на передних и по три – на задних, из чего следует, что жил он в густых зарослях, бегал по влажной почве и питался листвой и тонкими ветками.
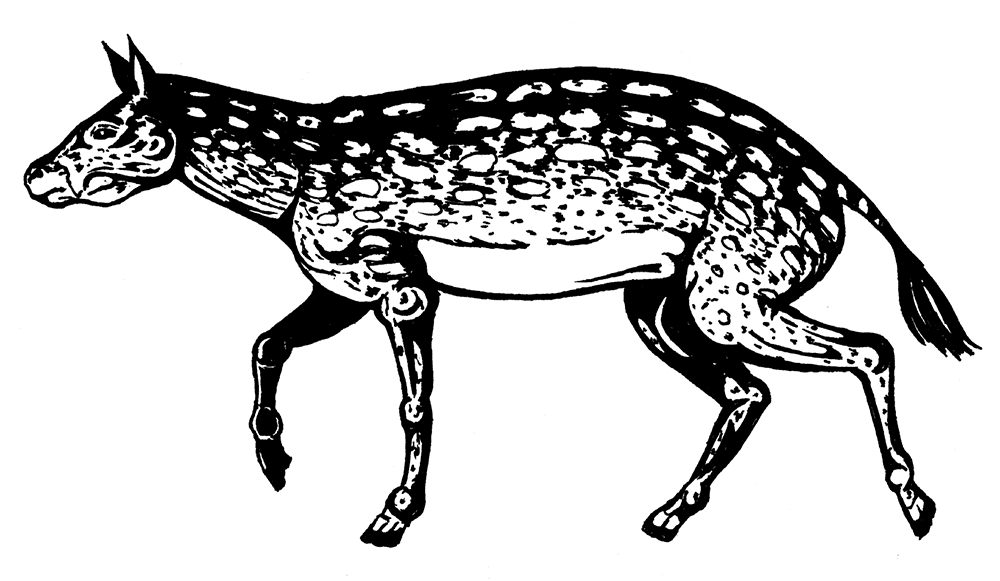
Eohippus angustidens
По мере изменения условий жизни исходная «кондиляртность» уменьшалась, а «лошадность» усиливалась, причём в нескольких эволюционных линиях независимо и немножко по-разному. Лошадиная ветвь представлена, например, последовательными Orohippus и Epihippus. У них всё ещё имелись по четыре пальца спереди и три сзади, а локтевая и лучевая кости предплечья оставались независимыми в память о роющих предках; по-прежнему низкокоронковые зубы помаленьку обзаводились гребнями для разрезания листьев; размеры тела помаленьку росли.
Маленькая тонкостьВ. О. Ковалевский на основе тщательнейшего изучения эволюции конечностей и зубов ископаемых лошадей и энтелодонтов пришёл к выводу, что, во-первых, разные части тела эволюционируют с неравными скоростями и, во-вторых, в случае обгона одной системы другой общая итоговая приспособленность может оказаться ниже, чем у конкурентов, у которых признаки менялись медленнее, но равномернее. В. О. Ковалевский разделил понятия инадаптивной эволюции – неравномерной и не вполне согласованной, накладывающей ограничения на последующие прогрессивные изменения, и адаптивной – более соразмерной, без крайностей и заскоков. Кроме того, В. О. Ковалевский первым в палеонтологии развил и последовательно применял морфофункциональный анализ – оценку эволюционных изменений с позиции их приспособительного смысла в данной конкретной экологической среде. Сейчас такой подход кажется само собой разумеющимся, но в XIX веке это был мощнейший прорыв; стоит ведь помнить, что эволюционная теория только-только была опубликована, а палеонтология возникала на глазах. Да и сейчас во многих палеонтологических трудах занудное перечисление морфологических отличий следует без малейшей попытки осознать их смысл.
В эоцене среди лошадеобразных выделились Eomoropidae, например Paleomoropus jepseni и Eomoropus из Восточной Азии и Северной Америки. Эти листоядные звери размером с овцу имели довольно крупные клыки в пасти и когти на пальцах и могут быть предками более поздних халикотериев. Некоторые подобные животные, например европейский Lophiaspis, иногда уже напрямую относятся к халикотериям; впрочем, эоморопиды могли быть и параллельно развивавшейся группой.
Примитивные парнокопытные Artiodactyla в раннем эоцене эволюционировали несколько медленнее, чем непарнокопытные, зато раскачались к концу эпохи. Первые представители – Dichobunidae в лице многих видов раннеэоценовых Diacodexis, среднеэоценовых Homacodon и прочих других – широчайше распространились по Европе, Северной Америке и Азии. Внешне они выглядели как карликовые оленьки с длинными тонкими хвостиками весом два-три килограмма; несмотря на принадлежность к парнокопытным, они имели ключицы и по пять пальцев с копытцами на каждой ножке. Подобны им, разве что крупнее и с короткими хвостиками, были и североамериканские Hypertragulidae. Удивительна рассинхронизированность эволюции передних и задних конечностей: у Hypertragulus всё ещё пятипалая когтистая кисть сочеталась с функционально двупалой стопой (сохранялись рудиментарные фаланги ещё одного пальца стопы, но совсем крошечные), а у Leptomeryx – четырёхпалая ручка с полностью двупалой ножкой. Показательны изменения зубов: верхние резцы уже успели исчезнуть, но у гипертрагулюса имелись довольно крупные клыки и острые премоляры, а нижние резцы были почти вертикальны, тогда как у лептомерикса клыки уменьшились, премоляры почти перестали отличаться от моляров, а нижние резцы направились горизонтально вперёд. Несмотря на примитивность, гипертрагулюсы дожили до миоцена.
Азиатские Archaeomeryx optatus и его родственники размером с зайца были и внешне похожи на зайцев: попрыгунчики с длинными задними и короткими передними ножками-шпильками, развитыми верхними резцами, но ещё и с клыками, и с длинным хвостом. Очень быстро, уже к концу эоцена, подобные существа доэволюционировали до семейства Tragulidae, примерами чего могут быть Archaeotragulus krabiensis и Krabitherium waileki из Таиланда.
Как вы там, потомки?Современные карликовые оленьки Tragulidae – азиатские Tragulus и Moschiola и африканские Hyemoschus aquaticus – просто напрашиваются на то, чтобы стать объектом загадок: саблезубый зверь, отлично плавает и даже ходит по дну, где ловит рыбу и раков, лазает по деревьям и лианам, день проводит в дупле, ночами выходит на поиски листьев, плодов, крыс, ящериц и всяческой падали. Кроме огромных клыков у самцов, оленьки обладают невероятно длинным языком и трёхкамерным желудком, а никаких намёков на рога у них нет. Весят они от полкило до восьми килограмм, африканские гиганты – до пятнадцати. По итогу, оленьки – едва ли не самые примитивные из нынешних парнокопытных, в их лице можно видеть осколок эоценового мира влажных тропических джунглей.
Саблезубость присуща и некоторым другим парнокопытным, например кабарге Moschus moschiferus, водяному оленю Hydropotes inermis и мунтжаку Muntiacus muntjak, причём у последнего клыки сочетаются с рогами.
Успешной и многочисленной группой эоценовых парнокопытных были Anthracotheriidae, например, древнейший мелкий Elomeryx и поздний двухсоткилограммовый Bothriodon (он же Ancodus); появились они, видимо, в Азии, потом оказались в Европе, а оттуда добрались и до Северной Америки в одну сторону и Африки – в другую. Особенно преуспели многочисленные виды Anthracotherium, дожившие от среднего эоцена аж до миоцена. Эти звери выглядели как приземистые тонконогие свино-тапиро-бегемотики с вытянутыми мордами, и ведь вправду они оказались предками гиппопотамов, а по менее вероятной версии – родственниками свиней. Набор зубов был практически полным, на передних лапках имелось по четыре пальца, а на задних – по пять. Такое строение оказалось практически универсальным для жизни в заросших болотах. Впрочем, антракотерии ещё не были слишком специализированы и могли уклоняться в разные стороны: некрупный бирманский Siamotherium pondaungensis бегал по открытым лесам и был вполне всеядным, не слишком уважая листья. Видимо, подобные антракотерии или очень родственные животные стали основой и для монстрических энтелодонов, и, возможно, для китов.

Anthracotherium
Как вы там, потомки?Современные копытные рождаются практически готовыми, работоспособными, иногда они даже почти сразу пытаются щипать траву, хотя, конечно, довольно долго ещё кормятся молоком. Повышенная самостоятельность резко повышает выживаемость, даже при небольшом числе детёнышей – у копытных обычно один на одни роды. Получается, что, с одной стороны, это высшая степень доделанности и прогресса на момент рождения среди млекопитающих, а с другой – парадоксальным образом копытные вернулись в идеал рептилий, у которых из яйца вылупляется уже готовая ящерка, только маленькая. По сравнению с примитивными млекопитающими, то есть нами, такая стратегия вроде как и продвинутая, но диалектически становится тормозом на пути развития разума, ведь если детёныш обеспечен с рождения всеми необходимыми программами поведения, то и обучение уже не нужно. Оттого мы – архаичны по сравнению с овцами, но мы – люди, а они – овцы.
Во второй половине эоцена часть парнокопытных Северной Америки стала увеличиваться в размере, превратившись в двухметровых кабаноподобных тварей Achaenodon – самых крупных парнокопытных в своих фаунах. Они были либо первыми и уже впечатляющими Entelodontidae, либо, вероятнее, гигантскими представителями Helohyidae. Ахенодоны сохранили весьма архаичную зубную систему, в том числе огромные толстые клыки, острые премоляры и бунодонтные – низкокоронковые бугорчатые – моляры, а их короткие массивные головы были чем-то средним между кондиляртровыми и свиными. Вероятно, эти монстры были не сугубо растительноядными, а хотя бы частично занимали нишу крупных падальщиков и могли грызть кости. Несколько позже их сместили с пьедестала Brachyhyops и Archaeotherium – уже несомненные представители энтелодонтов, расселившиеся и по Северной Америке, и по Азии, и по Европе, успешно перешедшие в олигоцен и давшие там великолепное потомство. Длинные челюсти археотериев были усажены примитивными зубами, в том числе огромными клыками, и снабжены крупными выростами для мощнейших жевательных мышц, о чём явно свидетельствуют и широкие скуловые дуги. Как и ахенодоны, археотерии, кроме растений, пожирали падаль; этому же способствовало и отличное обоняние, о чём мы можем судить по увеличенным обонятельным луковицам. Тяжеленная голова поддерживалась сильными мышцами, крепящимися на длинные остистые отростки грудных позвонков, отчего археотерии имели ссутуленный вид. Ноги, несмотря на сравнительно большую длину, не были приспособлены к быстрому бегу, а кисть и стопа адаптировались к ходьбе по влажному грунту. Эти твари стали конкурентами не только и столько растительноядным, сколько мезонихиям и креодонтам.
Отдельная ветвь парнокопытных – мозоленогие Tylopoda. Они настолько специфичны, что иногда выделяются в самостоятельный отряд. Называть их «парнокопытными» даже язык не поворачивается, ведь на пальцах у них не копыта, а толстые кривые когти, а кисть и стопа у современных – пальцеходящие, а не фалангоходящие; имеются у них и другие уникальные особенности, например овальные эритроциты, специфика зародышевых оболочек и строения желудка. Современные мозоленогие довольно однообразны – азиатские верблюды и южноамериканские ламы, а вот с эоцена по миоцен эта группа была многообещающа. Совсем не похожи на верблюдов низкорослые короткомордые Oreodonta (иногда определяются как Oreodontoidea, или Merycoidodontoidea), например североамериканские Protoreodon, Agriochoerus и Merycoidodon. По внешности это было что-то среднее между полутораметровой свиньёй и бегемотом, но с почти верблюжьими зубами и длинным тонким хвостом. Верхние клыки сочетались с клыкоподобным первым нижним премоляром, тогда как нижний клык был резцеподобным. Судя по специфической ямке на черепе впереди от глазницы, ореодонты могли иметь пахучие железы для мечения территории; о том же косвенно свидетельствует отличное развитие обонятельных частей мозга, например у Bathygenys reevesi. На кистях и стопах было по четыре длинных функциональных пальца и один маленький рудиментарный, все с когтями или копытцами; кости предплечья и голени не срастались. Несмотря на бочкообразность тела, коротконогость и общую примитивность конечностей, ореодонты были одними из первых жителей степей, где паслись их стада, состоящие из сотен животных, что видно по количеству находок. По факту ореодонты были едва ли не самыми массовыми животными своего времени. Надо думать, в последующем это благолепие истребили новые хищники и вытеснили более шустрые копытные.

Merycoidodon
Другие североамериканские мозоленогие – Oromerycidae, например Protylopus размером с зайца и Eotylopus величиной со среднюю антилопу, – сохраняли самый генерализованный облик и примитивнейшую зубную систему, зато в их предплечье локтевая срослась с лучевой, а в голени малая берцовая редуцировалась до невнятной шпильки. Эоценовые североамериканские Protoceratidae – мелкие Leptotragulus и Leptoreodon, а также более крупные Heteromeryx dispar и Pseudoprotoceras – были предками самых необычных рогатых верблюдов более поздних эпох; пока же они не слишком выделялись на общем фоне. Наконец, в эоцене появляются и настоящие верблюды Camelidae – североамериканские Poebrodon, известные лишь по нескольким зубам; будущее этой группы было впереди.
Совсем другим путём пошли родственники ореодонтов и верблюдов в Европе. Маленькие низенькие коротко-тонконогие и очень длиннохвостые Cebochoeridae, например Gervachoerus, сохраняли генерализованный кондиляртровый облик. У Choeropotamidae – Amphirhagatherium – хвостик укоротился, а задние ножки стали сильно длиннее передних, обеспечивая попрыгучесть; судя по бунодонтным зубам, они питались фруктами. Дальше всех пошли Cainotheriidae, например Cainotherium: от эоцена до миоцена эти крошечные – 15 см в высоту – копытные «зайчики» прыгали по зарослям и открытым степям. Несмотря на подвижность, кайнотерии сохраняли по четыре пальца на ручках и ножках. Кроликоподобным стало не только телосложение, но и череп, и зубы.
Серьёзнее выглядели европейские Anoplotheriidae, например Anoplotherium – очередные тапироподобные почти-мозоленогие с пятью толстыми пальцами на передних и четырьмя на задних коротких ножках, с длинной мордой и толстым кенгуруподобным хвостом. Предполагается, что аноплотерии могли вставать на задние ноги, вытягиваясь за высокорастущими листьями. Xiphodontidae в лице Xiphodon и его родни представлены, наоборот, длинноногими стройными двупалыми подобиями лам.
* * *Самые нестандартные парнокопытные среднего эоцена с самой нестандартной судьбой – Raoellidae в лице индийских Indohyus indirae и I. major. Эти мелкие зверюшки на тонких ножках оказываются ближайшими родственниками китов! Как ни странно, это вполне можно видеть на тварюшке меньше метра длиной. Но половину этой длины составляет мощный хвост, тонюсенькие косточки имеют чрезвычайно толстые стенки, а крошечная черепушка выглядит как арифметическое среднее между свиной и дельфиньей. Особенно же специфично утолщение медиальной стенки тимпанической пластинки височной кости при уменьшении толщины латеральной стенки: такая форма встречается только у китообразных, и случайное совпадение по столь особенному признаку маловероятно. Эти и другие особенности свидетельствуют об отличных способностях к нырянию; возможно, в воде индохиусы спасались от хищников, как это делают африканские карликовые оленьки, способные задерживать дыхание на четыре минуты. Судя по зубам, индохиусы были растительноядными или всеядными и точно отличались в этом отношении от самых первых китов. Более того, индохиусы явно не могли быть прямыми предками китов, ведь древнейшие китообразные уже существовали и даже найдены в тех же местонахождениях, но на примере этих тварюшек мы воочию видим «живых ископаемых» переходных форм.
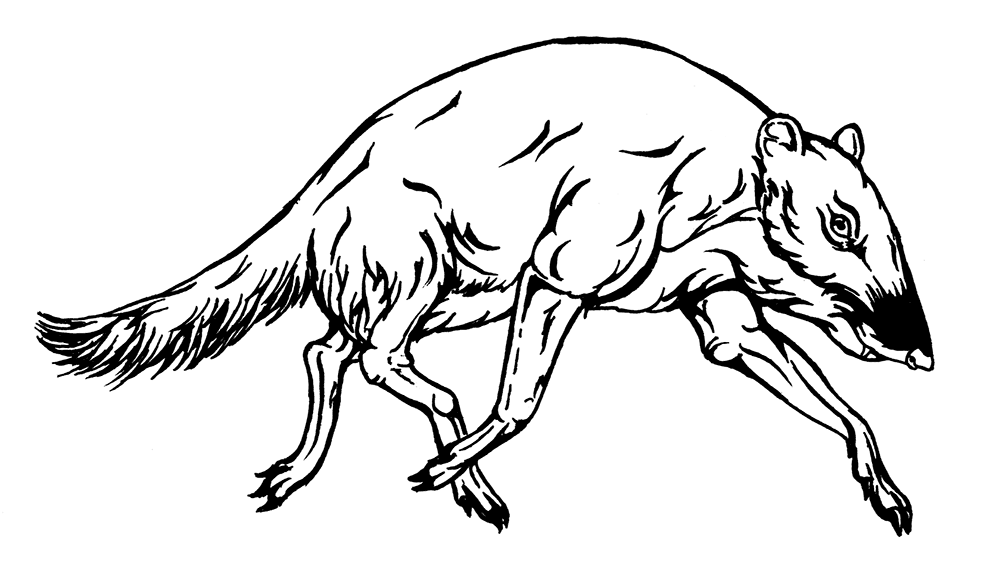
Indohyus indirae
Вообще, эволюция китообразных Cetacea – удивительнейшая тема. Начать с того, что Гималаи – родина китов. Казалось бы, как могли величайшие вершины мира породить самых водных зверей? Но в эоцене гор ещё не было, а их будущие заоблачные хребты были дном моря Тетис. А на берегах и мелководьях жили будущие киты. Кто конкретно из великого изобилия древнейших копытных был прямым предком – до сих пор вопрос. Генетические данные указывают на бегемотов как ближайших современных родственников, но бегемоты – это выжившая родня, они появились гораздо позже, да к тому же в Африке.
Одни из лучших кандидатов на роль предков китов – копытные хищники Mesonychia. В эоцене они расцвели пышным цветом, дав крайне харизматичных представителей: Mesonyx с приличного волка и такого же облика; масса видов гиеноподобных, но размером с медведя, Harpagolestes с широчайшим распространением от Северной Америки до Центральной Азии; Pachyaena, среди нескольких видов которой рекордных значений достигла P. gigantea – под сотню килограмм веса, а по смелым оценкам – даже до четырёхсот! В отличие от всех прочих хищников эоцена, мезонихии, будучи копытными, здорово умели бегать и, несмотря на традиционно тапироподобное тело, были хотя и не быстрыми, зато выносливыми. Также, мезонихии не развили специализированных хищнических зубов, их коренные зубы тупы, так что они могли быть не столько хищниками, сколько падальщиками.
Совсем особняком стоит Andrewsarchus mongoliensis из Монголии – крупнейший хищный зверь всех времён, со скуластым черепом 83 см длиной и 56 см шириной! На всякий случай, у белого медведя максимальная длина черепа 44 см, а у пещерного – 49 см. На таком фоне даже огромные клыки эндрюсархуса не кажутся огромными; а ведь клыкоподобным был ещё и второй резец, отделённый от обычного клыка маленьким третьим резцом. Одновременно моляры эндрюсархуса совсем тупые. Сочетание длинного зубного ряда без зазоров, широкой глотки и способности крайне широко открывать пасть говорит о всеядности зверя. Растопыренные вверх и в стороны глазки косвенно свидетельствуют об околоводной жизни. Проблема в том, что от эндрюсархуса сохранился лишь один-единственный череп, так что его истинный облик и образ жизни остаются под вопросом. Таксономическое положение эндрюсархуса загадочно, он родственен то ли мезонихиям, то ли энтелодонтам, то ли китам. Длинный приплюснутый крокодилообразный череп весьма напоминает черепа самых первых китов, так что эндрюсархус мог быть своеобразным сухопутным китом.
Другое дело, что в некоторый момент генетика показала родство китообразных и современных бегемотов, – и с этого момента многие специалисты резко поменяли своё мнение. Генетика – это же не скучная анатомия, гены не видать, стало быть, их анализ – это высшая степень магии, а потому даже многие прожжённые морфологи склонны доверять генетическим данным больше, чем собственным глазам. Крайне показательно, что в кладограммах, построенных на основе морфологии, мезонихии чаще оказываются ближайшей роднёй китов, но при включении в анализ молекулярных данных на первый план выходят бегемоты, а мезонихии удаляются в родню креодонтов и хищных. Но постойте! Ведь у нас нет вообще никаких сведений о генах мезонихий! Так может, дело не в родстве, а в тонкостях статистических методов и допущениях с предположениями? К тому же, строго говоря, одно не так уж исключает другое: черепа антракотериев, бегемотов, мезонихий и китов все так или иначе напоминают друг друга, так как восходят к кондиляртровому исходнику – низкие, плоские, с длинными вытянутыми челюстями, суженными спереди и широкими сзади, с огромными развёрнутыми скуловыми дугами, с тенденцией к увеличению клыков, с глазками, расположенными сбоку-сверху. И от такого неспецифического кондиляртрового корня легко выйти и на хищных мезонихий, и на растительноядных индохиусов с антракотериями и бегемотами, и на рыбоядных китов. В разных линиях сохранялись и усиливались разные черты, да к тому же добавлялись новые.
Маленькая тонкостьЕсть несколько версий, как и зачем киты вообще полезли в воду.
Если принять травоядную версию предков, то мелкие оленькоподобные зверьки могли просто прятаться под водой от злых хищников, а потом увлеклись нырянием. При мелких размерах, исходной клыкастости и недалёкости от насекомоядных исходников у них уже была некоторая склонность к мясоедству, примером чему служат современные карликовые оленьки, ловящие мелких животных, причём часто именно рыбу. И вот сегодня ты, трепеща, захлёбываешься на дне речки, спасаясь от кровожадного креодонта, рыщущего по берегу, а завтра сам с удовольствием лопаешь селёдку в океане.
Если же предками всё же были мезонихии, то они уже были хищниками и падальщиками. Такие твари могли собирать всякие вкусняшки на берегах водоёмов, а потом увлечься серебристой чешуёй – и уплыть за горизонт.
Если первые этапы окитения пока ещё скрываются в палеонтологическом тумане, последовавшая эволюция изучена просто отлично: известны все детали преобразования всех частей тела. Радостно наблюдать, как от одного вида к другому зверюшки меняются: стартовав 53–48 млн л. н., четвероногие собакоподобные ко времени 48–47 млн л. н. становятся всё более ластолапыми и тюленеподобными, к 42–37 млн л. н. – хорошо китообразными, но всё ещё с ножками, а около 34 млн л. н. – полноценными китами современного облика. Таким образом, на полное преображение понадобилось от десяти до двадцати миллионов лет – срок, вполне сопоставимый с появлением черепах из ящерицеподобных предков и прямоходящих разумных людей из совсем-совсем обезьян. Видимо, такой темп эволюции – более-менее норма, определяемая терпимым давлением отбора и скоростью появления удачных мутаций.
Древнейшие – ранне-среднеэоценовые – достоверные киты выделяются в семейство Pakicetidae; все они найдены на границе Пакистана и Индии. Сейчас их известно уже не так мало: Ichthyolestes pinfoldi размером с лису, Pakicetus attocki, P. inachus и другие виды – с собаку, Nalacetus ratimitus – с волка. У всех них отлично развиты и даже длинны все четыре ноги, кисть даже пятипалая, стопа – четырёхпалая. У них сохранялись ключица и третий вертел бедренной кости, а ведь они имелись лишь у самых ранних и примитивных парнокопытных. Шея была не то чтобы длинной, но нормальной. В черепе поражает мизерная заглазничная ширина, так что мозговая коробка соединялась с челюстями тоненьким стебельком, при огромных широко развёрнутых скуловых дугах. Вытянутые челюсти оборудованы гетеродонтными зубами, причём заклыковые зубы имеют выраженные бугорки; ноздри открывались на кончике морды. Почему же их относят к китам? Очень толстые кости приспособлены к нырянию; очень длинный и массивный хвост – залог будущего китовьего хвоста с лопастями; глаза расположены сбоку и сверху – опять же для ныряния; в слуховой капсуле медиальная стенка утолщена, как у китов, хотя полукружные каналы большие (у более поздних китов они малы), да и в остальном внутреннее ухо устроено скорее как у наземных животных, так что пакицеты неплохо слышали и в воде, и на суше. В итоге почти все особенности пакицетов промежуточны между наземным копытно-мезонихиевым состоянием и водно-китовьим. Судя по зубам, пакицеты уже могли быть рыбоядными, хотя по тем же зубам видно, что предки явно были растительно- и всеядными. Любопытно, что налацет найден в пресноводных речных отложениях, а не в морских, так что приспособление к нырянию и плаванию началось ещё до освоения морских побережий.