
The Open Sea: The World of Plankton
The microscopic plants must have some great advantage over larger plants in this floating drifting life. The smaller an object is the larger is its surface in relation to its volume. If we increase the size of an object—keeping its shape in the same form—the volume increases by the cube of linear measurement but the surface does so only by the square. This elementary fact is so important in the present discussion that it may be well to emphasise it by a simple concrete example. If we have eight small cubes of soap of the same size and press them together to form one big cube, the volume of this new cube will then be eight times that of one of the smaller ones, whereas its surface will be only four times as large. We have of course lost all the surfaces that were pressed and fused together. Inversely, the more we cut up our soap into smaller and smaller cubes, the more surface will each new cube have in proportion to its volume. A cube the size of one of our little plants will have a surface-volume ratio many hundreds of times as large as that of a cube with a side no more than an inch or two. A large surface-volume ratio is a great advantage to our little plants in at least two important respects. Firstly, the larger the surface in relation to mass the greater will be the frictional resistance to the water which will retard its sinking and so enable it to remain more easily in the upper sunlit layers. Secondly, since absorption must take place through the surface, the larger its surface in proportion to its volume the more readily will it be able to take up for its needs enough of the necessary mineral salts which may be present in the water in only very small amounts. This indeed may be the cardinal factor which has prohibited the development of larger plants in the plankton; but for this they might well have evolved bladder-like floats to support their larger mass, as some animals have done. Each tiny plant, as a single cell, can also take better advantage of the scattered sunlight than can a number of such cells massed together.
It is at first difficult to believe that these finely scattered and microscopic plants can really form a vegetation which has sufficient bulk to support all the teeming animal life of the sea: the dense populations of planktonic crustaceans, the vast shoals offish and all the invertebrate animals on the sea-bed. Yet we know this must be so. Some estimates of the actual quantities of plants present in a cubic metre of sea-water will be given later in the chapter; here, in passing, we will only note that, given suitable conditions, the amount of plant life produced under a given area of sea may well exceed that produced for the same area in a tropical forest. Just as our coal supplies are giving us the energy stored up in the great primaeval forests of some hundred million years ago—so is the energy in the petrol, which drives our motor and flying age, derived from that originally trapped from the sunlight by the tiny planktonic plants in the seas of long ago. According to current geological theory, the great supplies of mineral oil have been formed, in the course of ages, from the remains of marine organisms buried in sedementation under specially favourable conditions which are not yet fully understood. It is most likely that the planktonic crustaceans, whose modern representatives are so rich in oil, would in the past be the main contributors to the supplies of petrol we are burning up today; those crustaceans, of course, derived their energy either directly or indirectly from such tiny plants as we are now considering.
A microscope of sufficient power to enable us to see a great deal of this world of planktonic plants and animals need not be an elaborate one, nor need it cost much more than a good pair of field-glasses. We shall want some glass slides and coverslips, small dishes (such as watch-glasses), pipettes, i.e. old-fashioned fountain-pen fillers, for picking up very small plankton animals, and some glass jam jars; apart from that, all we need is a tow-net and line with which to collect the plankton from a rowing boat or any larger vessel that can be made to go slowly enough.
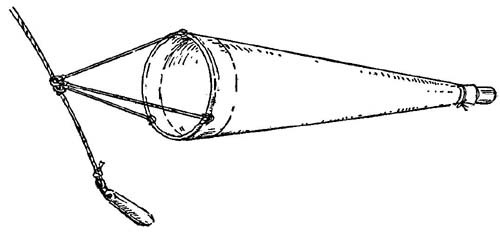
FIG. 12
A simple form of the plankton collecting tow-net.
A tow-net can be bought from the laboratory of the Marine Biological Association at Plymouth (address: The Laboratory, Citadel Hill, Plymouth) or it can be home-made. It consists essentially of three parts: in front is a hoop made either of light galvanised iron or strong cane and provided with three bridles of cord which will come together at a small ring or shackle for attaching to the towing rope; next comes the actual net, a conical bag made of a fabric which will act as a fine sieve; lastly at the end of the net is a small collecting jar, either a glass honey-jar or one made of zinc or copper with a slight lip. Such a simple tow-net is shown in Fig. 12. For ordinary collecting purposes a hoop of 18 inches diameter will be quite sufficient. The net is best made of the silk ‘bolting cloth’ used by millers for sieving flour, but a good quality muslin will do if this cannot be obtained; with a mouth of 18 inches it should be almost five feet in length. If it is to be homemade great care should be taken in cutting out the material in order to ensure that a perfect cone is formed; if lop-sided it will not fish properly. It is a good plan to pin together a paper model to serve as a pattern; this will also enable one to see how best to use the material with as little waste as possible. Round its wide mouth a canvas or calico band is sewn for attachment to the hoop; it may either be provided with a series of eyes for lashing it on or it may be folded over the hoop and sewn to. enclose it, leaving gaps where the towing bridles are secured. At the hind end is sewn another canvas band to form a small cylinder, say 2½ inches in diameter, which will slip closely over the mouth of the collecting jar and be firmly held in position by a tightly tied tape. It is well to be provided with two such tow-nets; one made of the very finest material for the collection of the small plants—the finest bolting-cloth has 200 threads to the linear inch—and one of coarser material, having about 60 threads to the inch, for the capture of the somewhat larger animals. The coarser net lets most of the plants go through its mesh but filters a very much larger quantity of water more quickly and so captures the larger more active animals which are only rarely taken in the finer net.
To collect the phytoplankton the fine net should be towed just a little way below the surface. A weight, say a 71b. lead, is slung at the end of the rope and the net attached a little way above it. The essence of successful tow-netting is to tow very slowly, never at more than 1½ knots. If it is towed faster the water will not be filtered quick enough; the net will just push a mass of water in front of it which will prevent any more water entering it. A ten minutes’ tow may give quite a large enough sample. Most of the plankton will have passed down into the jar at the end as it is towed; a number of specimens, however, may still be sticking to the inside of the net as it is taken from the water, so that it should be carefully washed down from the outside with water from a bucket, to flush them into the jar.
Our sample will contain a vast number of both plants and animals. In this chapter we will concern ourselves only with the former, which are so small that they must be looked for with the compound microscope. After bringing our sample home and letting it stand for a little we should take only a few drops at a time with a fine pipette from near the bottom and place them on a slide under a coverslip; now we shall hunt with the low-power lens and then turn on the high-power to examine each new specimen we find. We shall not, of course, expect to find examples of all the different kinds in one sample but there may well be representatives of several of the more important groups. The most prominent members of the phytoplankton are the diatoms. They are unicellular algae differing from all other algae in having a cell wall which forms a siliceous external skeleton enclosing the cell like a glass box. The pigment bodies, or chloroplasts, which enable the plant to make use of the energy of sunlight are not the usual bright green of chlorophyll but a brown or brownish-green pigment closely allied to it. The siliceous skeleton is in two parts which fit together like the top and bottom of a pill-box; indeed some of the diatoms are just like a pill-box in form, but many others are drawn out into all manner of fantastic shapes. When first we see a sample of plankton rich in diatoms under the high power of the microscope it is like looking at a group of crystal caskets filled with jewels as the strands of sparkling protoplasm and groups of amber chloroplasts catch the light. Every plant or animal cell consists of a mass of protoplasm with a more or less central body, the nucleus, which appears to govern its life; it is characteristic of the diatoms that, in addition, the protoplasm usually has large cavities in it containing clear fluid. The nucleus is usually central and surrounded by a mass of protoplasm; radiating from this and forming an irregular network are protoplasmic strands stretching across the cavities like the spokes of a wheel to join up with a layer of protoplasm which lines the inner surface of the boxlike covering. More rarely, in some forms, the nucleus may be in the layer of protoplasm at the side. The pigment granules usually lie more or less regularly spaced against the cell-wall, where they are exposed to the light; if, however, the light is too intense, they come close together either down the strands to the centre or to some other part of the cell where they can partly screen one another from the harmful effects of the rays.
The top, bottom and sides of the glass-like box are not made of just plain sheets of silica; their surfaces are sculptured with all manner of striations, pits and perforations forming intricate patterns peculiar to the different species. This detail of design has always made the diatoms favourite specimens with microscopists, not only on account of their beauty, but because they are such excellent objects with which to test and display the quality of their instruments in the higher ranges of magnification. They are now being put under the electron microscope which can give a micrograph with a magnification of up to 100,000 times; this has at once revealed an arrangement of structure far more elaborate than that seen with the highest powers available in the optical systems (Hendy, Cushing and Ripley, 1954). Instead of there being just one system of pits or perforations in their walls, some forms are shown to have smaller and yet smaller ones, secondary and tertiary systems, on inner layers of silica; in others the wall is more like a basket of spiral threads intricately woven together. Some of the perforations measured were less than a ten-thousandth of a millimetre in diameter and the surrounding walls were of equal thickness. These delicate lattice systems have at least two important qualities for floating plants: they give strength with lightness and at the same time provide a framework for presenting a greatly increased surface area of protoplasm to the surrounding water.
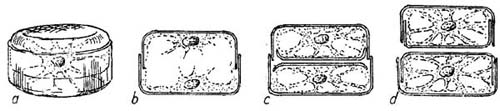
FIG. 13
Diagrams showing the division of a simple pillbox-like type of diatom. a, a sketch of the cell before division has begun, b to d sections through the diatom after cell division to show stages in the formation of the new skeletal cell walls. Note that the upper cell in d is smaller than the original cell b.
Diatoms normally reproduce by simply dividing in two. The nucleus divides first and then the protoplasm becomes separated into two masses, each containing a nucleus, one at either end of the box; each mass of protoplasm now forms, between it and the other mass, a new valve as the halves of the pill-box are called. These new valves each fit closely their own part of the old box; we have in fact two pillboxes now instead of one, as is shown above in Fig. 13. They may separate entirely, or in some species they may remain attached to form long chains. It will be realised that in this process of repeated division by forming new half-boxes within the old, the average size of the diatoms so produced will tend to get smaller and smaller; at each division, as shown in the drawing, one of the new boxes will be the same size as the old one but the other must be smaller. Thus we find a considerable range in the size of diatoms of the same species, but there must be a limit to this reduction. After a certain number of such divisions there is formed what is called an auxospore, by which the original size is recovered; throwing off the old valves the cell becomes a bladder-like mass of protoplasm within which new valves are formed two or three times the size of the old discarded ones. Some diatoms seem to do this at definite seasons whereas others do so only at intervals of two or three years. By taking sample measurements of the diatoms forming some of the dense concentrations which are carried by currents about the North Sea, planktologists have been able to identify individual patches in their wanderings by the regular decrease in the size of their component cells (Wimpenny 1936; Lucas and Stubbings 1948). Some of these concentrations, as we shall see in Chapter 15 have been thought to have a marked effect upon the herring fisheries; their wanderings may therefore be of economic interest.
In addition to forming auxospores, diatoms may produce what are called resting spores when conditions become adverse. The contents of the cell become concentrated in a central mass which forms a new thick wall of a different but characteristic shape and the old cell-wall is discarded. They now either sink into the deeper water layers or right to the bottom where they will remain till more suitable conditions return; thus some may pass the winter in a resting state and then come up to start active life again in the spring. Planktonic diatoms are not definitely known to have any sexual phase, but in some a number of smaller spores (microspores) have occasionally been observed to be formed within the cell-wall and it has been thought that these may be gametes (sex cells), but this is not yet established.
Not all diatoms are planktonic; in the shallow coastal regions there are numbers living on the bottom where sufficient light reaches it; these have much thicker shells than the more delicate floating forms and are more uniform in character. What makes the planktonic diatoms so interesting is the variety of devices that have been evolved to assist in their flotation. It is not the purpose of this book to attempt a systematic treatment of the groups of animals and plants. Here we shall just refer to some of the more important kinds of diatoms in relation to their mode of suspension. Dr. Marie Lebour’s excellent book (1930) on the planktonic forms should be studied for a full account.
Although so very small they have a considerable range in size; a few exceptionally large ones may be over one millimetre in diameter and the smallest may be but a few thousandths of this. Sketches of examples of the different genera to be mentioned are shown in Fig. 14 and Plate 1, and some are also included in the photographs in Plate I. A number have the typical pill-box form, such as members of the genus Coscinodiscus and some of these are of comparatively large size; a few like C. concinnus may be just visible to the naked eye. These kinds have large vacuoles and would seem to be buoyed up by globules of oil. Members of this genus live singly, but others of allied genera may remain after division attached together to form long chains; the cells of Paralia and Guinardia are linked rigidly together by their valve surfaces, those of Thalassiosira form flexible chains as they are strung together by fine threads of protoplasm, and others like Lauderia are still more loosely held together by irregular strands of slime. The cells of Thalassiosira also produce such slime-strands but for a different purpose—around the margin of each valve (i.e. each ‘pill-box lid’) are a number of small hollow spines from which can be extruded long slender threads of slime so that they radiate on all sides like the strands of thistle-down and indeed act in the same way to assist in parachute-like support.
The remainder of the planktonic diatoms, while essentially built on this pillbox plan, have each valve or half-box modified into all sorts of shapes which increase their surface area in relation to their volume and so give greater frictional resistance to sinking. Some are flattened like thin sheets of paper and often twisted to some extent; these usually remain attached together to form long ribbons: such are Bellarochia, Eucampia and Streptotheca. Others are drawn out either into long thin hair-like forms such as Thalassiothrix longissima or into the more rigid pencil or needle-like members of the genus Rhizosolenia, pointed at each end; in the former the division plane between the two valves runs lengthwise along the thread whereas in the latter it occurs transversely to the long axis. Other forms again, such as Biddulphia and Corethron, increase their surface area by being provided with spines. This last method is developed to a remarkable extent by the many species of the genus Chaetoceros whose cells have four very long hair-like processes (two extending from each valve); long chains of these cells are formed and held together by their curving processes becoming interlocked with those of adjacent cells close against their point of origin.
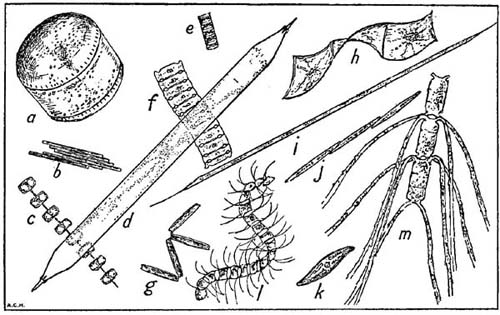
FIG. 14
Some characteristic plankton diatoms not shown in Plate I, all magnified ×90 diam. a, Coscinodiscus concinnus; b, Bacillaria paradoxa; c, Thalassiosira gravida; d, Rhizosolenia styliformis; e, Paralia sulcata; f, Bellarochia maleus; g, Thalassiothrix nitzschioides; h, Streptotheca thamensis; i, Rhizosolenia hebetata (form semispina); j, Nitzschia seriata; k, Gyrosigma sp.; l, Chaetoceros curvisetus; m, Ch. convolutus. The actual length represented by the longer side of this figure is 1/20th of an inch.
There are many species of Biddulphia in our waters, but one, B. sinensis (Plate 1) is of special interest; it is now one of our commonest diatoms, often occurring in dense concentrations, yet it was unknown in European waters before 1903 when it was first recorded in the Heligoland Bight. It is a well known inhabitant of the coastal waters of the Indo-Pacific region, extending from the Red Sea to the coasts of China. It seems likely that it must have been brought, perhaps in ballast water, by some ship to the mouth of the Elbe; being tolerant of wide ranges of temperature and salinity, it found our waters congenial and spread rapidly. In the following years it was recorded further and further to the north until it reached a point a long way up the Norwegian coast where its further spread was probably checked by too cold water. More extraordinary, in view of the prevailing currents, was its spread down the Channel and into the Irish Sea; in the same year, 1909, it was reported for the first time both at Plymouth and off Port Erin in the Isle of Man. This would appear to provide evidence of an occasional reversal of the usual current flow up the Channel; indeed such a reversal has been suspected by some oceanographers on other grounds. Or was it transported to the western Channel and Irish Sea in the same way as it had apparently reached the Elbe? We may never know the answer to that. Some have maintained that Biddulphia sinensis must have been a native of our waters all the time and only just noticed at the beginning of the century; this, however, can hardly be so because of extensive collections that were made, particularly by the Kiel planktologists, throughout the eighteen-nineties.
In coastal regions we may find a number of typical bottom-living diatoms carried up into the plankton, such as the boat-shaped members of the genus Navicula. They are capable of a remarkable gliding movement, thought to be produced by a flow of protoplasm passed out through a slit in the wall of the cell. Gyrosigma is another bottom form often met with in the shallow-water plankton. Other related forms such as Bacillaria and Nitzschia are more planktonic but typical of coastal waters. B. paradoxa forms bands of long slender cells held together side by side like the planks in a raft yet each capable of sliding up and down along its neighbours; N. seriata, in contrast, forms long strings of narrow boat-shaped cells end to end with just their tips overlapping and in contact.
It is impossible to mention all the kinds of diatoms of our seas in such a general review, and I shall only refer to one other species, one which may well attract attention: Asterionella japonica (Plate 1). Its cells are rod-like but thickened at one end; by these thickened ends the cells remain attached to one another to form beautiful radiating star-like clusters.
In striking contrast to the brown-green colouring of the diatoms is the brilliant green sphere of Halosphaera viridis which may reach a size of nearly a millimetre in diameter; it and one or two closely allied species are the only representatives in the marine plankton of the Yellow-green Algae or Heterokontae. H. viridis is found over the whole Atlantic from the tropics to the far northern branches of the North Atlantic current off Spitsbergen. In autumn it is often brought into the northern North Sea in large numbers and is usually found floating very near the surface. It is exceptional in its mode of reproduction; it does not divide in two, but when full-grown undergoes multiple fission into a large number of small spores which break out of the surrounding envelope and swim, like the flagellates about to be described, by the use of whip-like locomotory organs. The full life-history has not yet been observed; whether after fusing with others or not, these spores must eventually give rise to the little green spheres which gradually grow to a full size again.
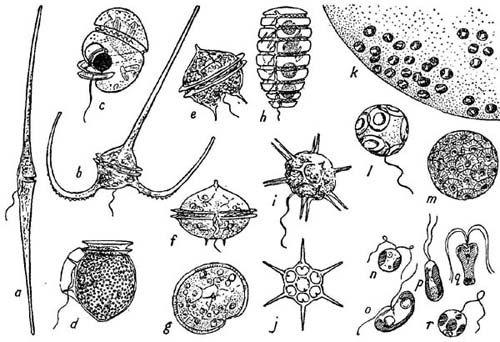
FIG. 15
Some flagellates of the plankton. a–h, Dinoflagellates: a, Ceratium fusus (×180); b, C.macroceros (×200); c, Protoerythropsis vigilans (×320) (note clear spherical lens against dark eye-spot); d, Dinophysis acuta (×400); e, Peridinium granii (×360); f and g, P. ovatum (×320), side and top view; h, Polykrikos schwarzi (×250); i and j, the Silicoflagellate Distephanus speculum living and half of skeleton (×320); k, a very small part of the large gelatinous capsule formed by the tiny cells of Phaeocystis; l and m, Coccolithophores (×1000): Coccolithus huxleyei and Coccosphaera leptopora; n–r, some of the smallest flagellates (×1500): n, Dicrateria inornata; o, Hemiselmis rufescens; p, Isochrysis galbana; q, Pyramimonas grossii; r, Chromulina pleiades. Original drawings except c from Marshall (1925), h from Lebour (1925), m from Murray and Blackman (1898) and n to r from Parke (1949).
All the remainder of the planktonic plants belong to the big assemblage of organisms known as flagellates of which there are many different kinds. A selection of the commoner forms is shown in Fig. 15. They are all characterised by possessing at least one, and often two, of the motile whip-like processes termed flagella, with which they draw or propel themselves through the water and are thus able to keep up in the sunlit surface layers. These flagellates are claimed for study by both botanists and zoologists, for among them are indeed both plants and animals—and some which have the characters of both in one. Some possess green pigments allied to chlorophyll or even chorophyll itself, and so feed as true plants; others lack pigment and may feed either by absorbing organic substances through their surface or actually live as animals by capturing particulate food; yet again, others may combine the methods of plant and animal feeding. In this lowly group of organisms the animal kingdom has not yet become fully separated from the plant kingdom. However, most of the planktonic flagellates are in fact plants and most of them have a small red ‘eye-spot’ or stigma which is sensitive to light and so enables them to tell whether they are moving towards or away from the radiant energy necessary to build up their food. If you are able to obtain a plankton sample very rich in these small green flagellates you will be able to see how readily they are attracted upwards towards the light. Fill a tall narrow glass jar with the sample and cover the lower three-quarters with thick brown paper. Now if you leave it for half an hour in the full light of the window you will find on removing the paper that the top quarter of the jar is distinctly greener than the rest; the little flagellates from the whole jar have become concentrated in the sunlit zone. By standing a sample of sea water in the light you may be able to grow a more abundant culture of these little flagellates and so give a more striking demonstration of this experiment. There are some of the Dinoflagellates (members of the family Pouchetiidae) which have a much more elaborate light-sensory organ furnished with both a lens and a pigment-cup; indeed it might almost be called an eye. One of these is shown in Fig. 15c.