
Flowers of the Coast
Generally speaking, all succession is directed towards developing the most complex vegetation which the climate will permit, no matter what the nature of the original habitat may have been. The ultimate vegetation produced in this way is called the climax formation or the climatic climax. The communities making up this formation will be more or less stable and will not be seriously threatened by new invaders. In most of England and the southern part of Scotland, if the vegetation were left completely undisturbed, oak or beech forest would eventually be developed. In the north of Scotland and most of the central portion also, if we exclude the tops of the higher mountains, the climatic climax would, however, be pine forest, an association of the Northern Coniferous Forest Formation. In comparatively recent times, most of the British Isles was forested in this way, but the large-scale felling of our woodlands during the Middle Ages and later has almost obliterated the natural forests. Nowadays, as a result of intensive agricultural operations, the climax formation is rarely reached in the course of natural succession. Where suitable areas exist, which are not cultivated or grazed, the absence of suitable seed-parents in the immediate neighbourhood precludes the development of natural woodland. Ecologists recognise, however, a number of relatively stable subclimaxes in the vegetation of this country, which are developed under the conditions which are normally present.
Any natural succession of communities which replace each other in a particular habitat is called a sere. Thus those which succeed each other in a salt-marsh all belong to the halosere, salt being the master-factor controlling each of them, and those developing on blown sand to the psammosere (Greek: psammos=sand). The sea-coast provides practically the only habitats in this country where one can see a more or less complete series of communities starting with bare ground and finishing with a type of vegetation which remains comparatively stable under the particular conditions. Elsewhere, succession can be most easily observed in an area which has previously carried some fairly stable type of vegetation, but which has subsequently been modified in some way or other. This is well illustrated when a wood is felled or a heath is burnt and is known as secondary succession. Good examples of this type of development can also be seen along the coast, as for instance when the surface vegetation on a sand-dune is broken through and the strong winds produce a “blow-out” (see here), which is then recolonised in much the same way as the fresh sand on the newest dunes (Pl. XXI). As another example, the seaward edge of a salt-marsh sometimes becomes eroded as a result of a sudden change of current or for some other reason. The original vegetation is thus destroyed, but in course of time the mud on which it originally grew may be colonised once more to form what is called “secondary marsh,” usually at a different level from the original one.
When we come to look into the reasons why particular plants grow where they do, we find that there are a large number of factors to take into account. Most of these are closely inter-related in the effects they produce, but it is worth while to discuss briefly some of those which are especially important in determining coastal vegetation.
The climate of the country is obviously of the greatest importance, for it controls such factors as the duration and intensity of the sunlight, the range of temperature, the rainfall, the humidity of the atmosphere and the strength of the winds. Climatic factors show their effect most clearly when vegetation is studied on a broad geographical basis, but even in a relatively small area like that of the British Isles the effects of small differences in climate are quite noticeable. Thus the average rainfall and the humidity of the air is much greater on the west coast than on the east, and this probably accounts for many small differences in the distribution of plants along the two coasts. It is certainly responsible for the much richer moss flora found on the western sand-dunes compared with those on the east coast, and may partly account for the occurrence of certain typical “Atlantic” species along our western and south-western coasts. In the same way, the mean temperature in the North is distinctly lower than that in the South, and this is one of the factors responsible for the absence or rarity of a number of plants in Scotland and northern England, which are comparatively common in the South, and also for the fact that certain characteristic north European plants are only found in the North.
Wind is obviously a very important factor in all coastal habitats. Its most pronounced effect is that it increases the loss of water vapour from the leaves of plants by constantly bringing dry air into contact with them. As a result, the growth of many seaside plants is considerably retarded and they are often found in a very stunted form. To combat this, many coastal plants adopt a mat or rosette habit for much of the year. Exposed parts of the coast are generally destitute of trees, and such few trees as do occur near the coast are usually found tucked away in sheltered valleys, or combes as they are called in the West Country. Trees and hedges in coastal areas often assume very distorted forms, which show clearly the direction of the prevailing winds (Pl. VIII). This is due to the fact that only the shoots on the leeward side can develop normally, those continually exposed to the prevailing winds being dried off and killed. In this way they appear to have been blown over by the strength of the wind, whereas actually their peculiar shapes are due to the unequal development of the shoots on their two sides. The effect of wind in retarding growth is most marked on the east and north-east coasts, which are exposed to the driest winds, although it is very noticeable on any of our coasts.
Another group of factors to be considered depend upon the general topography of the habitat and may be called physiographical factors. The angle at which the ground slopes, the aspect or direction of the slope and the height of the land above sea-level, are examples of these. The familiar coastal processes of erosion, silting and the blowing of sand, which are discussed in Chapter 2, also come into this class. In addition, the prevalence of strong winds along the coast, whose effects have just been described, is clearly due to a combination of climate and topography. It is hardly necessary to give illustrations of the result on the vegetation produced by all these factors; the relation of the highly specialised community of plants which are found on mobile sand with their habitat, for instance, is sufficiently obvious. Some of them, however, become particularly important when we consider cliff-vegetation. Thus the angle at which the cliffs slope largely controls the amount of soil available for supporting plants in the rock crevices, and will indeed determine the stability of the surface of the cliff itself, if it is composed of soft material. The height above the sea will also determine the amount of spray to which the habitat is exposed, and most cliffs show some zoning of the vegetation which can be correlated with this factor. The direction towards which a cliff faces is also important in determining the amount it will be exposed to the prevailing winds and thus, indirectly, the amount of spray it is likely to receive, and will also control the duration of the periods of shade. There is often a marked difference in the vegetation of cliffs with different aspects, in particular those on the opposite sides of small islands.
Another group of factors, which in some ways show the most pronounced effects on the composition of the vegetation, are those related to the physical and chemical properties of the soil. These are called edaphic factors (Greek: edaphos=the ground). On the coast the commonest physical characteristic of most habitats is that of a poor water-supply. Sand-dunes, shingle beaches, and most cliffs are all subject to periodical drought conditions, which are aggravated by the drying winds. We shall see in the next chapter that the leaves of many seaside plants are equipped with devices of various kinds to check excessive loss of water, and that their root-systems are often very extensive. The amount of air contained in the soil is also related to its physical state, and it is noticeable that a number of plants, such as marram-grass on dunes, sea purslane in salt-marshes, and the shrubby seablite (Suaeda fruticosa) on shingle grow luxuriantly only when their roots are well aerated.
The chemical nature of the soil is also of great importance. Salt is obviously the master-factor in determining the highly specialised vegetation of salt-marshes, and the presence of halophytes in other coastal habitats, such as shingle beaches and exposed cliffs, shows that there also salt spray is deposited in sufficient amount to be an important factor. The ultimate vegetation developed on sand-dunes also varies greatly with the amount of calcium carbonate initially present in the sand. We have already seen that, if this is small, it will be washed out of the surface-layers in time, and that typical plants of acidic soils like heather and heath may eventually appear, as the supply of humus increases. Many west coast dunes, however, have been formed from sand which contains so much calcium carbonate in the form of broken shells that the relatively slow leaching action of the rain has produced little effect on it. As a result, the final vegetation on these dunes remains fundamentally calcicole (lime-loving), and is remarkably similar in composition to the grassland commonly found on chalk and limestone. In the same way, chalk and limestone cliffs may be expected to show some different plants from those which are found on acidic rocks.
Finally, we must say a word about the effects on the habitat caused by living organisms. These are called biotic factors (Greek: bios=life), and include the activities of man and his animals, the effects of rabbits, birds and insects and those produced by the plants themselves. The effects of previous generations of plants in altering the physical and chemical properties of the soil have already been briefly discussed. As far as man is concerned, his activities are less in evidence along the coast than in most parts of the country, since coastal areas do not lend themselves well to agricultural development. Nevertheless, in a thickly populated area like ours, there is no region where the hand of man has not played some part in modifying the vegetation. For instance, large areas of many salt-marshes are used for the grazing of cattle, which has the effect of restricting some plants but not others. Many old salt-marsh areas, too, have been completely transformed by drainage operations or the construction of sea-walls to exclude the tides, and the laying out of golf-courses has altered the vegetation in sand-dune areas in a number of places. Moreover, in certain districts marram-grass has actually been planted to stabilise shifting sand-dunes, and elsewhere rice-grass has been employed in a similar way for reclaiming salt-marshes, so that it is often impossible to distinguish between natural and partly artificial vegetation. Nor should it be forgotten that the large-scale felling of the native forests all over the country in the past has had the indirect effect of preventing the natural development of the climax vegetation in many suitably undisturbed areas along the coast.
Rabbits are frequently responsible for considerable modification of the vegetation, and are often extremely common in coastal areas. In particular, the grassland on the tops of cliffs is often infested with them, and the older sand-dunes provide a veritable rabbit’s paradise. In all probability, the somewhat stunted vegetation which is so characteristic of such areas results as much from its being continually nibbled by rabbits as from its exposure to strong winds. Some plants, however, are more attractive to rabbits than others, so the actual composition of the vegetation may be considerably altered. Even salt-marshes are not exempt from the attentions of rabbits; in some districts, for instance, it is unusual to see more than a quite small proportion of the sea-aster plants reaching the flowering stage. Birds also sometimes have a marked effect on the vegetation, particularly when large colonies gather on small islands for breeding purposes. Needless to say much excreta is deposited on the cliff-ledges and cliff-tops near their nesting sites, and the increase in the amount of nitrogen and phosphates in the soil produced in this way has the effect of altering the composition of the vegetation considerably.
The above brief summary can do no more than suggest the kind of factors which must be looked for if we are to make any attempt to understand why coastal vegetation is distributed as it is, and why particular species occur in some places and not in others. Our knowledge of these matters is still extremely incomplete, and it is well to realise that much valuable information can still be easily collected by amateur botanists who are prepared to make a fairly detailed survey of the vegetation in a particular habitat and to keep their eyes open for the factors which have been responsible for its composition.
CHAPTER 4 FORM AND HABIT OF COASTAL PLANTS

THE MAJORITY of the plants we find growing round the coast have to contend with unusually harsh conditions, and many of them are specially adapted to enable them to survive in their inhospitable habitats. In this chapter we shall consider some of the characteristic growth-forms they adopt.
Undoubtedly the main problem for most of these plants is to obtain adequate supplies of water, particularly in the early stages of their growth. This applies both to those growing in such obviously dry habitats as sand-dunes, shingle beaches or rocky cliffs, and to those growing in saline ground, such as salt-marshes or brackish swamps, although the reason for the difficulty is quite different in the two cases. The whole question of water-supply is sufficiently fundamental to merit discussion in some detail.
To deal first with the dry habitats; the whole trouble here is that they do not retain sufficient quantities of water in their surface layers, since the “soil” they provide is largely made up of coarse particles. The water-holding power of a soil depends in the first instance on the size of its particles. If these are large, water can percolate easily through them, and will also evaporate more quickly because of the large air-spaces between them. Thus the greater the number of small particles, the longer the soil will take to become dry after rain. Furthermore, it is a well-known fact that water tends to stick on to the outside of all relatively small particles on account of the force known as “surface-tension,” and since the total surface-area of a given weight of small particles is clearly greater than that of the same weight of coarse particles, the finer the soil the greater its powers of retaining water. But in addition to a lack of small particles, there is usually a shortage of humus in all the habitats in question. This important material, consisting of dead plant-remains in the process of decay, has already been briefly referred to (see here). Without discussing the varied forms in which this organic matter can occur, the amount present in a soil can be roughly guessed from its colour. Thus dark-coloured “peaty” soils contain the greatest amount and sands the least. All farmers are familiar with the fact that adequate quantities of this material are necessary in all “light” (i.e. coarse) soils, if they are not to suffer from frequent drought conditions. Humus possesses great powers of absorbing water, chiefly because much of it is usually in the form of very small particles of what are called “colloidal” size (i.e. they are so small that they easily pass through a filter-paper, and take a long time to settle when they are suspended in water). Quite apart from this, it is a valuable source of plant food, partly on account of the nitrogen it contains, but principally because it absorbs valuable salts and prevents them from being washed away.
The plants growing in saline habitats also have trouble with their water-supply, though of a very different kind. Here there is often an abundance of water, but it is, of course, salt water. As a result, the plants may suffer from what has been called a “physiological drought.” This means that, despite an abundance of water in the soil, they are unable to make use of it on account of the high concentration of salt it contains. Many salt-marsh plants, therefore, live under conditions of partial drought rather similar to those encountered in other coastal habitats. As evidence of this, it can often be noticed that they are greatly benefited by the dilution of their soil-water, when a spell of wet weather occurs in the summer. Indeed many, though not all, halophytes can grow luxuriantly in ordinary garden soil.
Another characteristic of coastal habitats is that they are all to a greater or lesser extent exposed to strong winds. The most important result of this, as has already been pointed out, is to increase the rate of evaporation of water at the leaves (transpiration). This causes plants to draw further on their slender water-supplies, and if these are inadequate, wilting may take place. Thus wind aggravates the results of the water-shortage.
In order to understand the various ways in which maritime plants deal with this fundamental problem of water-supply, it is necessary to say a word about two processes, common to all plants, which are specially important in this connection. These are osmosis and transpiration.
OSMOSIS
All plants obtain the water and soluble salts required for their growth through the agency of cells situated near the ends of their roots, which are known as the root-hairs. These cells are filled with sap, which contains small quantities of soluble salts and much larger amounts of soluble organic substances, such as sugars, in solution.
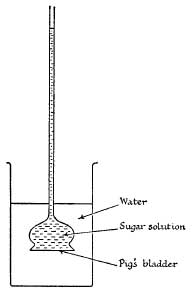
FIG. 2.—Experiment to demonstrate osmosis.
When the root-hairs are in close contact with the soil-water, a suction pressure is developed through the walls of these cells, called the osmotic pressure. As a result of this, water passes into the cell and temporarily dilutes the sap. This is a familiar chemical phenomenon and can easily be demonstrated in a number of ways. If any solution is enclosed in what is called a “semipermeable membrane” (i.e. one which will allow water, but not dissolved substances, to pass freely through it), the osmotic pressure of the solution will cause water to be sucked into the solution through the membrane.
A simple way in which this can be demonstrated is to tie a piece of pig’s bladder very firmly over the end of a small funnel (preferably a “thistle funnel”). If the funnel is now filled with a strong solution of (say) cane sugar and is immersed in a vessel containing water, the water will soon start to pass through the membrane into the sugar solution and rise up the stem of the funnel (Fig. 2). If a long piece of glass tubing is attached to the stem, it will be noticed, after a day or two, that a column of water several feet high has risen up the tube. It is important to realise that the height of this column is not a measure of the pressure exerted by the initial sugar solution, since this solution is naturally becoming steadily more dilute as the water enters it. In point of fact the osmotic pressure of a 15 per cent solution of sugar is about 10 atmospheres, or sufficient to support a column of water of well over 300 feet in height!
The walls of the root-hair cells function in exactly the same way as semipermeable membranes, though they allow considerable amounts of the substances dissolved in the root-water to pass through them also. Measurements of the osmotic pressures exerted by the cell-sap of many different plants have been carried out. These have been found to vary considerably, but a value of about 10 atmospheres for normal plants (mesophytes) can be taken as an average figure. As a result of this large suction pressure, it is obvious that a root-hair which is freely supplied with water will soon become distended with diluted sap and develop a corresponding balancing pressure. This is called the “turgor pressure.” An equilibrium between this and the osmotic pressure, resulting in the cessation of the flow of water into the cell, would soon be reached were it not for the fact that water is continually passing from the root-hairs into the root and stem of the plant. It is this movement which maintains a flow of water through the whole plant, the excess water being eliminated largely through the leaves. The process by which water is conducted through a plant is extremely complex, and there is nothing to be gained by attempting to discuss it here. The important thing to understand is that, provided water is in contact with the root-hairs, a steady flow into the root can be maintained. On the other hand, if insufficient water is available in the soil, the plant may not be able to obtain an adequate supply to build up its turgor pressure, with the result that the whole plant becomes limp and is said to “wilt.” Obviously this danger is greater if water is eliminated too rapidly by the leaves, and plants growing in dry habitats are often provided with devices to prevent excessive transpiration.
Measurements of the osmotic pressures exerted by both halophytes and xerophytes have been shown to be, in general, much larger than those characteristic of normal plants. We have mentioned 10 atmospheres as a typical value for mesophytes, whereas 40 atmospheres would appear to be an average value for plants in the former classes. Indeed, some desert plants have been shown to exert pressures running up to the enormous figure of 100 atmospheres and more. Obviously this greatly increased power of suction must be of much assistance to plants growing in arid soils in enabling them to obtain what little water there is. In the case of halophytes, a high osmotic pressure is virtually essential if they are to overcome the considerable pressure of the salt water in which they have to grow. Ordinary sea-water has an osmotic pressure of about 20 atmospheres, but in a salt-marsh the concentration of salt may become very much higher during a spell of dry weather in those areas which are not submerged by every tide. If halophytes were incapable of exerting a greater osmotic pressure than that of the salt water in a marsh, osmosis would take place in the wrong direction and water would be sucked out of the plant into the soil-water. Thus the plant would not only fail to obtain its water-supply, but would lose much of the water it already contained. This effect can easily be demonstrated by putting any normal plant into salt water, when it will be seen to wilt in a very short time.
A good deal of work has been done on the measurement of the osmotic pressures developed by halophytes when growing in salt solutions of varying concentrations. The results show clearly that they fluctuate considerably and are able to alter rapidly to adjust themselves to changes in the concentration of salt in the soil-water. This adaptability accounts for the wide tolerance shown by many halophytes growing in different parts of salt-marshes. The whole problem of the mechanism by which water is absorbed by plants is very complex, and the above account is much simplified in order to explain the main principles.
TRANSPIRATION
Passing now to the other end of the plant, we must say something about the process by which the surplus water is disposed of at the leaves. This is known as transpiration. On the surface of any leaf a number of minute pore-like openings are to be found which are called the “stomata.” Each stoma usually takes the form of a slit between two elongated cells known as “guard cells,” lying side by side (Fig. 3(c).) The opening or closing of the pore is controlled by the swelling or contraction of this pair of cells. Thus, when the turgor pressure of the plant is high, the cell-walls expand and the slit is opened to aid the elimination of water. When the water-supply is less abundant, the turgor pressure falls and the cells contract so that they lie with their walls in contact with each other, thus closing the slit. It should be emphasised that the stomata are not only concerned with the elimination of water-vapour but are also the organs through which the plant absorbs carbon dioxide and gives out oxygen in the carbon assimilation process (photosynthesis). They are in fact the openings through which the exchange of all gases takes place, although to some extent the whole surface of the leaf and even the stem functions in this capacity. When the external covering or “cuticle” of the leaf is thick, however, the process is largely confined to the stomata. Usually these occur more thickly on the under-surface of the leaf, as being better protected from the drying influence of the sun. Only in water-plants with floating leaves are they confined to the top surface. Although the number of stomata found on the leaves of different plants varies greatly, there do not appear to be any fewer on those belonging to halophytes or xerophytes than on the leaves of normal plants. A moderately large leaf with an average density of stomata may possess several millions of such openings.