
Клеточные и молекулярные механизмы патогенеза иммуновоспалительных ревматических заболеваний
Эта фаза носит характер диффузных или очаговых мононуклеарно-клеточных инфильтратов и формирования гранулём различной степени зрелости, которые могут носить гигантоклеточный характер. Гранулёмы формируются начиная с этапа фибриноидных изменений и характеризуются накоплением активированных макрофагов и веерообразным расположением этих клеток вокруг масс фибриноида и фагоцитозом продуктов дезорганизации основного вещества соединительной ткани. Наиболее явственно описанная картина встречается при ревматической лихорадке, но базисные характеристики представлены в той или иной мере и форме при всех других ревматических заболеваниях. Патоморфологическая идентификация гранулём при ревматических заболеваниях является свидетельством участия гиперчувствительности замедленного типа (ГЗТ) как клеточного типа иммуновоспалительных реакций. Фаза клеточных реакций является фазой реализации иммунопатогенетического аспекта ревматических заболеваний. На светооптическом уровне эта фаза выражается в виде макрофагальной реакции, фагоцитозе клеточного и тканевого детрита, с последующей презентацией антигенного материала лимфоцитам (функции АПК), атрофии лимфоидной ткани с замещением лимфоидных элементов плазматическими клетками (плазматизация лимфоидной ткани), миелоидной метаплазии центров размножения фолликулов лимфатических узлов и селезёнки, формированием фолликулоподобных структур, периваскулярной инфильтрации мононуклеарами (лимфоциты, моноциты), что формирует хронический продуктивный панваскулит.
Завершающий этап указанных изменений – развитие процессов фиброза и склероза, при которых ключевое значение приобретает активность фибробластов. Важной характеристикой этого процесса является сочетание сформировавщихся очагов фиброза и склероза с признаками мукоидного набухания и фибриноидных изменений, что свидетельствует о непрерывном, прогрессирующем течении заболевания. Необходимо подчеркнуть, что все изменения, возникающие при коллагенозах в полной мере относятся и к стенкам сосудов, что обуславливает формирование васкулитов. Описанные процессы свойственны таким хроническим системным иммуновоспалительным ревматическим заболеваниям, как ревматическая лихорадка (РЛ), ревматоидный артрит (РА), системная красная волчанка (СКВ), системная склеродермия (ССД), дермато-полимиозиты (ПМ), системные васкулиты (СВ).
Какие же молекулярно-клеточные процессы лежат в основе формирования иммуновоспалительного клеточного инфильтрата при ИВРЗ?
1.1. Клеточный инфильтрат при иммуновоспалительных ревматических заболеваниях
Морфологическим субстратом ХПВ является КВИ. С позиций общей патологии в формировании КВИ при ИВРЗ ключевая роль принадлежит состоянию гиперреактивности системы соединительной ткани. Ещё И. В. Давыдовский писал: “гиперергические реакции различной сложности наблюдаются часто и в процессе аутоиммунизации“ и далее “таким образом, воспалительный процесс в принципе является иммунизаторным” [4].

Ипполит Васильевич Давыдовский (1887–1968), советский патологоанатом, академик АМН СССР. Крупнейший философ-медик XX века. С его именем связано становление и развитие патологической анатомии в нашей стране
В ситуации иммуновоспалительного процесса при ревматических заболеваниях гомеостатическая регуляция матрикс-клеточных взаимодействий существенно нарушается. Формируются сложные каскады межклеточных взаимодействий, из которых наиболее патогенетически значимые – это триада: макрофаг-лимфоцит-фибробласт, в которой связующим элементом являются макрофаги (Мф). Мф и гистогенетически близким им клеткам отводится центральная роль в формировании КВИ. Эта роль обусловлена тем, что эволюционно выработанная основное функциональное предназначение Мф, а именно – утилизация отжившего клеточного и тканевого детрита, а также индукция адаптивного АГ-специфического иммунного ответа, в контексте ИВРЗ, трансформируется в презентацию ауто-АГ CD4+ и CD8+Т-лимфоцитам и аутоиммунным ответом на продукты дезорганизации рыхлой волокнистой неоформленной соединительной ткани [97]. Накопление Мф в очаге воспаления является признаком разгара этого патологического процесса.
КВИ является динамической структурой, отражающей этапность, рецидивирующее течение и исход ИВРЗ. КВИ может принимать организованные и неорганизованные формы. К первым относятся фолликулоподобные структуры и ГЗТ-гранулёмы, ко вторым – диффузный клеточный воспалительный инфильтрат. Исход указанных форм КВИ может быть в виде его исчезновения; в виде образования рубца вследствие несбалансированного фиброза; в виде обострения воспаления in situ за счёт притока лимфоцитов, моноцитов, нейтрофилов и других клеток; в виде персистенции воспаления.
В процессах несбалансированного фиброза видное участие принимают активированные Мф. Участие этих клеток выражается в том, что продукция ими фибронектина усиливают аттракцию фибробластов в зоне воспаления и стимулируют их коллагенсинтетическую активность. Кроме этого важными факторами активации и пролиферации фибробластов является фактор роста фибробластов 2 (FGF2) и тромбоцитарные факторы роста (PDGF-A, – B, – C и – D) [40]. Под влиянием этих и других факторов микроокружения фибробласты могут нарабатывать коллаген, но также способны секретировать и коллагеназу, вызываюшую деградацию основного вещества соединительной ткани, т. е. имеет место сочетание явлений фиброгенеза и фиброклазии, обуславливающих несбалансированный фиброз. Отметим роль макрофагальных ферментов. Протеазы макрофагов способны расщеплять любые белковые компоненты матрикса соединительной ткани, влияя на аутоантигенность последних. Однако в зонах прогрессирующего фиброза количество макрофагов, продуцирующих коллагеназу, существенно уменьшается.
Воздействие кандидатных триггеров иммуновоспалительных процессов приводят к первоначальным признакам дезорганизации рыхлой волокнистой неоформленной соединительной ткани, связанной с деполимеризацией основного вещества соединительной ткани (протеогликаны), разволокнением коллагенового каркаса, под воздействием в т. ч. коллагеназ, эластаз и гиалуронидаз, что сопровождается явлением метахромазии, повышением гидрофильности ткани и мукоидным набуханием, повышением сосудистой проницаемости и развитием воспалительного отёка. Нарушение структуры коллагеновых и эластических волокон, увеличение уровня фибронектина, продукция провоспалительных хемокинов и факторов роста эндотелиоцитов способствуют усилению воспаления и ангиогенеза [145].
Патоморфология диффузного КВИ на светооптическом уровне при всех ИВРЗ характеризуется определённой унифицированностью клеточного состава. Воспалительный инфильтрат является преимущественно мононуклеарным. В количественном отношении доминирующими клетками в составе КВИ являются лимфоциты, затем идут клетки макрофагально-моноцитарного ряда.
На рис. 1 представлена типичная картина ХПВ при СКВ и при ПМ. Видно, что инфильтрат и при СКВ в дерме, и в поперечнополосатых мышцах при ПМ располагается компактно, довольно выраженный и является мононуклеарным.

Рис. 1. Клеточный воспалительный инфильтрат в дерме при СКВ, слева (Х100). Такой же инфильтрат в поперечнополосатых мышцах при полимиозите, справа (Х100), по материалам [5]
Организованная форма инфильтрата в виде гранулёмы, или узелка, представлена на рис. 2 слева. В данном случае определяется гранулёма в миокарде при РЛ. Видно, что и в этом случае инфильтрат является мононуклеарным, компактным, морфологически идентифицируемым как узелок. Всем ИВРЗ характерно развитие продуктивного васкулита той или иной степени выраженности с периваскулярной локализацией КВИ. На рис. 2 справа представлена картина мононуклеарной инфильтрации при узелковом полиартериите. Важной особенностью этого препарата является наличие периваскулярного фибриноидного некроза. Но это явление определяется не всегда. На рис. 3 справа представлена картина продуктивного васкулита с активным периваскулярным инфильтратом, но без признаков некроза. Одновременное присутствие признаков некроза с последующей организацией этих зон в склеротические и активного мононуклеарного инфильтрата свидетельствует о рецидивирующем характере воспаления при ИВРЗ.
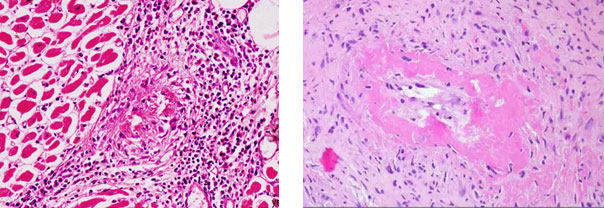
Рис. 2. Узелковый продуктивный миокардит при ревматической лихорадке, слева (Х100). Справа картина узелкового полиартериита, виден периваскулярный некроз и преимущественно мононуклеарная инфильтрация (Х100), по материалам [5]
Первыми клетками, реагирующими на воздействие кандидатных триггеров ИВРЗ, являются клетки макрофагально-моноцитарного ряда, дендритные клетки (ДК) и активированные эндотелиоциты. Фагоцитарная активность этих клеток по отношению в т. ч. к продуктам дезорганизации соединительной ткани является базисной в отношении индукции межклеточной кооперации по типу “фагоциты-лимфоциты” и формирования микроокружения, оптимального для индукции аутоиммунных процессов in situ. Фагоцитоз Мф указанных продуктов, а также материала аутологичных клеток, подвергшихся некробиозу и некрозу in situ, ограниченный протеолиз фагоцитированного материала обеспечивает включение клеточно-молекулярного механизма антигенной презентации Т-лимфоцитам, в изобилии находящихся в составе воспалительного инфильтрата. В очаге ХПВ идут интенсивные процессы неоангиогенеза. Не исключается, что активированные эндотелиоциты могут приобретать в т. ч. и АГ-презентирующие свойства.
В этих условиях наблюдается известный патоморфологический признак аутоиммунных процессов – плазматизация лимфоидной ткани. В-лимфоциты подвергаются дальнейшему созреванию и превращению в плазматические клетки (CD38+) – продуценты ауто-АТ. Одновременно, активированные В-лимфоциты, находящиеся в микроокружении продуктивного воспаления могут выполнять функции АГ-презентирующих клеток (АПК), внося тем самым свой вклад в прогрессирование аутоиммунного процесса.
Важным является феномен эктопической экспрессии молекул МНС класса I и II на Мф и ДК при ИВРЗ, индуцирующих презентацию ауто-АГ аутоспецифическим клонам Т-лимфоцитов и последующее развитие аутоиммунного ответа [77].
В составе КВИ иммуногистохимически идентифицируются Т-лимфоциты (CD3+), Т-фолликулярные хелперные клетки (Tfh CD4+клетки), Т-цитотоксические клетки (CD8+), В-лимфоциты (CD20+), плазматические клетки (CD38+), макрофаги (CD68+), фолликуллярные дендритные клетки (фДК), в меньшем количестве нейтрофилы (Нф), эозинофилы, тучные клетки [42,87,89]. Согласно нашим данным с составе КВИ при ревматоидных синовитах и при дерматомиозите иммуногистохимически идентифицируются преимущественно Т-клетки (CD3+, CD4+), В-клетки (CD20+), макрофаги (CD68+). Также отмечалась тенденция к периваскулярной локализации КВИ и высокий уровень экспрессии ингибитора апоптоза – онкопротеина bcl-2 и маркёра ангиогенеза – CD31 [10,11]. Субпопуляционный состав КВИ имеет дифференциально-диагностическоле значение. Так при полимиозите CD8+T-клетки являются преобладающими в КВИ, тогда как CD4+T-клетки преимущественно проникают в мышечные волокна при дерматомиозите [98].
Необходимо отметить, что активированные мононуклеары непосредственно влияют на активность фибробластов. Эти клетки способны не только стимулировать функции фибробластов (участников КВИ), но и тормозить их, выступая в качестве истинных регуляторов фиброгенеза in situ. Так IFN-γ, продуцируемый Th1 CD4+лимфоцитами, макрофагальный IFN-β, а также TNF-α, способны тормозить коллаген-синтетическую функцию фибробластов [52,170]. Фенотип клеток в составе КВИ коррелирует с эффективностью лечения, что показано, в частности, при РА [48].
Резидентные Мф, а также Мф, трансформировавшиеся из моноцитов гематогенного происхождения, гетерогенны и разделяются на две субпопуляции – М1 и М2. М1 макрофаги, экспрессирующие мембранные маркёры CD215, CD80 и CD86, являются активными участниками воспаления, продуцируя провоспалительные и другие цитокины (TNF-α, TNF-β, IL-6, IL-1β, CCL2, IL-8, IL-12, IL-23, IFN I типа, GM-CSF), способствуют прогрессированию этого процесса, в то время как М2 макрофаги, имеющие фенотип CD206, CD209, продуцируют противовоспалительные цитокины (TNF-β, IL-10, IL-4, IL-13), обладают противовоспалительными эффектами и вносят свой вклад в процессы ремоделирования соединительной ткани и репаративной регенерации. Возможны взаимные переходы этих клеток. Поляризация М1/М2 зависит от хемо- и цитокинового окружения и наличия рецепторов к ним [99, 166].
Прямое участие клеток макрофагально-моноцитарного ряда в индукции аутоиммунного ответа продемонстрировано при СКВ. Показано, что моноциты гематогенного происхождения в составе КВИ способны дифференцироваться в плазмоцитоидные дендритные клетки (пДК), экспрессирующие высокий уровень CD86 и продуцирующие IFN-α. Такого рода пДК способны активно презентировать ауто-АГ аутореактивным Т-клеткам [25, 47].
Важный вклад в индукцию аутоиммунного ответа вносит т. н. “дефектный” фагоцитоз, при котором фагоцитированный материал может служить источником ауто-АГ. Основное предназначение фагоцитарной активности – утилизация клеточного и тканевого детрита – при ИВРЗ приобретает новые качественные особенности, трансформирующие АГ-презентирующую функцию Мф в направлении аутоиммунизации. Эти процессы документированы, в частности, при СКВ [65].
Неэффективное очищение макрофагами места воспаления от фрагментов клеток, подвергшихся апоптозу, также может служить триггером аутоиммунных процессов. При этом количество Мф (CD68+) в составе КВИ может существенно превышать нормативные показатели, что показано при системной склеродермии [66,97]. Важно, что при этом заболевании М2 макрофаги способствуют активации резидентных фибробластов и прогрессии фиброза за счёт продукции трансформирующего фактора роста β (TGF-β), фактора роста эндотелиоцитов (VEGF) и тромбоцитарного ростового фактора (PDGF) [29].
“Дефектный” фагоцитоз показан и при синдроме Шегрена. При этом синдроме фагоцитоз клеток, подвергшихся апоптозу, сопровождается индукцией аутоиммунного ответа, увеличением уровня провоспалительных цитокинов (IL-6, IFN-γ, TNF-α, IL-1β), IL-18, хемокинов (CXCL8, CXCL10), VEGF и PDGF [64].
Инфильтрация Мф синовиальной оболочки является одним из важных признаков РА. Патогенетическое значение активированных Мф в составе КВИ при РА определяется способностью этих клеток продуцировать избыточное количество провоспалительных цитокинов, из них ключевой – это TNF-α, а также IL-1, IL-6 и IL-12. Иммуногистохимическими методами показано присутствие этих цитокинов в синовии при РА и определена корреляция воспалительного процесса с их уровнем in situ [171].
Активация Мф связана с экспрессией поверхностной молекулы CD40, имеющей свойства рецептора. Взаимодействие CD40 с растворимыми и мембран-ассоциированными молекулами CD154 индуцирует продукцию Мф ряда металлопротеиназ (MMP-1, MMP-3, MMP-9, MMP-11, MMP-12), а также IFN-γ-индуцированную продукцию оксида азота. Молекулы CD154, принадлежащие к семейству TNF, в комбинации с IFN-γ могут вызвать переход М2 Мф в М1 Мф. Таким образом, взаимодействие CD40/CD154 способствует прогрессированию продуктивного воспаления. Источником CD154 являются активированные CD4+клетки, фибробласты и тромбоциты [105, 169].
Активированные макрофаги/моноциты продуцируют хемокины (СС-хемокины), IL-8, моноцитарный хемоаттрактант (МСР-1), макрофагальный воспалительный протеин (MIP-1α), провоспалительные цитокины – IL-1, IL-6, TNF-α и иммунорегуляторные цитокины – IL-10, IL-12. Эти растворимые факторы являются патогенетически важными при ХПВ [130].
Представленный спектр растворимых макрофагальных факторов, их специфичность и многофункциональность позволяет привлечь в место формирования КВИ лимфоциты, моноциты, нейтрофилы и модулировать их активность в соответствии с конкретными условиями микроокружения. Эффекторные Th1 CD4+лимфоциты мигрируют в очаг воспаления и, продуцируя IFN-γ, IL-2, TNF-α, обеспечивают развитие ГЗТ. На этом этапе возможно взаимодействие этих клеток с макрофагами, фагоцитировавших продукты дезорганизации основного вещества соединительной ткани. Это взаимодействие опосредуется через молекулу CD40 на поверхности макрофага, а также с участием IFN-γ.
Клетки в составе КВИ могут подвергаться некротическим и некробиотическим изменениям. Подобное повреждение клеток сопровождается появлением эндогенных сигналов опасности, называемых аларминами, или DAMPs, активирующих фолликулярные дендритные клетки (фДК) и плазмацитоидные дендритные клетки (пДК). DAMP способны взаимодействовать с рецепторами врождённого иммунитета (TLR-2, – 4, – 7, – 9) на ДК и активировать их. Роль ДК в активации адаптивного иммунитета, ведущего к выработке ауто-АТ, хорошо известна. В эктопических зародышевых центрах при ИВРЗ DAMP – активированные ДК, секретируя провоспалительный цитокиновый каскад, прежде всего IFN I типа, индуцируют процессы продуктивного воспаления, в т. ч. и иммунный ответ на ауто-АТ. Указанные явления определены при РА, при СКВ и при синдроме Шегрена в слюнных железах [125].
К числу наиболее актуальных эндогенных молекул-аларминов, имеющих несомненное патогенетическое значение при ИВРС, относится высокомобильный групповой белок 1, обозначаемый как HMGB1. HMGB1 является негистоновым ядерным белком, освобождающемся из клеток, подвергшихся инфекционному повреждению, некрозу, некробиозу, клеточному стрессу. Взаимодействуя с рецепторами врождённого иммунитета (TLR-2, – 4, – 7, – 9), экспрессирующихся на ДК и Мф, HMGB1 способствует продукции IFN-α, IL-1β, TNF-α плазмацитоидными ДК и Мф, тем самым способствуя прогрессированию продуктивного воспаления in situ и усилению АГ-презентирующей функции ДК и Мф. Кроме этого, имеются данные о том, что HMGB1 способен индуцировать анти-HMGB1-антитела, которые относят к общему классу анти-ядерных ауто-АТ при ревматических заболеваниях [120, 167].
Немаловажным свойством HMGB1 является также способность этого белка активировать тканевые металлопротеиназы (ММР1–9) и тканевой плазминоген, активность которых вносит существенный вклад в дезорганизацию рыхлой волокнистой соединительной ткани. Напомним, что к классу ММР принадлежат ферменты коллагеназы и эластазы. Указанные процессы документированы при СКВ [23], при РА [158], при синдроме Шегрена [51], при полимиозите [162].
На рис. 3 представлено фото экспрессии HMGB1 при коллаген-индуцированном артрите. Видно, что иммуногистохимически позитивная реакция на HMGB1 определяется на большинстве клеток в составе КВИ.

Рис. 3. Экспрессия HMGB1 при коллаген-индуцированном артрите, слева (Х100), по материалам [120]. Продуктивный васкулит, периваскулярная инфильтрация мононуклеарами, справа (Х100), по материалам [5]
При интерпретации патогенетической значимости КВИ при ревматических заболеваниях в настоящее время внимание уделяется новому феномену, называемому “внеклеточными ловушками”. Первоначально этот феномен был отнесён к сфере антиинфекционного иммунитета, впоследствии появились данные, не всегда однозначные, об участии “внеклеточных ловушек” в аутоиммунных процессах, в т. ч. и при ревматических заболеваниях – СКВ, РА, васкулитах [76, 83]. В случаях, когда этот процесс осуществляется клетками макрофагально-моноцитарного ряда “внеклеточные ловушки”, обозначаются как “метозис”. В случаях, когда этот процесс осуществляется Нф, он обозначается как “нетозис”. Суть этого феномена сводится к уникальной серии внутриклеточных событий, посредством которых ядерное содержимое, включая хроматин, смешивается с клеточными белками и затем вытесняется из клеток с образованием внеклеточных структур, способных “захватить” и убить микроорганизмы [32,50].
Процессы нетозиса и метозиса относят к формам программируемой клеточной гибели. Первые публикации касательно нейтрофильных “внеклеточных ловушек” (NET) появилось в 2004 году и за этим быстро последовало описание этого феномена и в отношении других клеток – макрофагов, моноцитов, эозинофилов, базофилов, тучных клеток. Поскольку процесс метозиса имеет некоторые черты сходства с фагоцитарным актом, то была предложена точка зрения, согласно которой при метозисе могут присутствовать процессы внутриклеточного ограниченного протеолиза, связанные с презентацией АГ детерминант. В пользу подобного взгляда выступают данные, свидетельствующие о цитрулинизации ДНК-связанных белков – гистонов в процессе нетозиса и приобретения ими ауто-АГ свойств, в частности, при РА [79].
При СКВ функцию нетозиса берёт на себя популяция гранулоцитов низкой плотности (LDG), которая отсутствует у здоровых людей. Процесс LDG-нетозиса при СКВ сопровождается появлением ауто-АГ детерминант, способствующих дальнейшему повреждению соединительной ткани, последующей активации пДК и гиперпродукции IFN I типа. Также предполагается, что при СКВ нетозис является неким связывающим звеном между появлением аутоантител при дезорганизации основного вещества соединительной ткани и индукции процесса метозиса, осуществляемыми пДК и выработкой ауто-АТ против ДНК [35, 57].
Клеточные компоненты нетозиса могут активировать В-клетки через BCR-клеточный рецептор и TLR9 рецептор, с последующей продукцией аутоантител против ДНК и кателицидина LL-37 [92]. Показано, что активированные эндотелиоциты также способны прямо стимулировать Нф к нетозису, которые, пребывая в этом состоянии, могут уже сами вызвать повреждение эндотелия [61].
Как видно из представленных результатов исследований, формирование КВИ при ИВРЗ – это многокомпонентный процесс, направление развития которого зависит от степени и характера дезорганизации рыхлой волокнистой соединительной ткани, микроокружения, клеточного состава и функциональной активности КВИ, спектра продукции и рецепции провоспалительных цито- и хемокинов и других растворимых факторов. Организация КВИ подвергается определённым закономерностям и характеризуется формированием морфологических форм, имеющих важное патогенетическое значение при ИВРЗ.
1.2. Эктопические лимфоидные структуры, или эктопический лимфоидный неогенез
Уникальной особенностью исходов КВИ является возможность приобретения клеточным инфильтратом определённых структурированных форм, отражающих в определённой мере нозологическую специфичность ревматических заболеваний и потенциальную возможность индукции аутоиммунного ответа в locus morbi. Одновременно эти процессы сочетаются с разностадийностью хронического воспаления, когда в воспалённой ткани определяются признаки мукоидного набухания и фибриноидных изменений с явлениями склероза и фиброза. Морфологическая трансформация КВИ в структуры, напоминающие вторичные лимфоидные органы (лимфатические узлы, пейеровы бляшки, селезёнка) получила название “эктопические лимфоидные структуры” (ELS) или “эктопический лимфоидный неогенез”. ELS приобретают многие особенности лимфоидных фолликулов вторичных лимфоидных органов, включая компартментализацию зон, богатых Т-клетками и В-клетками, и накопление фолликулярных дендритных клеток (фДК). Наиболее ярко ELS представлены при ревматоидных синовитах в виде фолликулоподобных структур в субсиновиальном слое [9].
На рис. 4 в качестве сравнительной иллюстрации представлены фото герминативного центра в интактной нёбной миндалине слева и герминативный центр фолликулоподобной структуры в синовиальной оболочке при РА справа. Видно, что в общей морфологической организации этих двух структур прослеживается значительная аналогия.
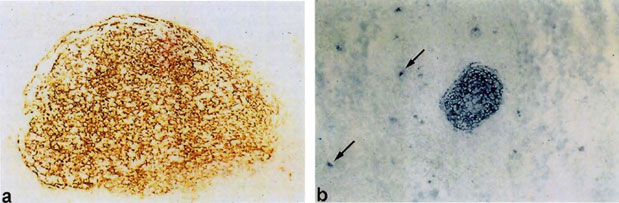
Рис. 4. Слева (а) представлен герминативный центр в интактной нёбной миндалине, справа (b) аналогичная структура в синовиальной оболочке при РА (Х 100), по материалам [88]
Клеточное микроокружение в ELS, широкий спектр продуцируемых и рецептируемых клетками воспалительного инфильтрата провоспалительных цито- и хемокинов, создают оптимальные условия для индукции аутоиммунного ответа in situ. CD4+ T-лимфоциты, находящиеся в составе КВИ, являются ключевыми клетками в процессах активации В-лимфоцитов, их дифференцировки в плазматические клетки и продукции последними ауто-АТ. Этот факт отражает известный патоморфологический признак иммуновоспалительных ревматических заболеваний, а именно – плазматизация лимфоидной ткани. В условиях ELS функции Т-хелперных клеток среди всей субпопуляции CD4+клеток берут на себя фолликулярные Т-хелперные клетки (Tfh), несущие фенотип CD4+ CXCR5+ Bcl6+. Tfh-клетки продуцируют важный иммунорегуляторный фактор IL-21 и хемокин CXCL13. IL-21 – это цитокин, который способствует пролиферации В-клеток в зародышевых центрах (GC) и их дифференцировке в плазматические клетки. ИЛ-21 могут продуцировать и активированные макрофаги в условиях ELS [146].