
Палеонтология антрополога. Три эры под одной обложкой
Гнётовые Gnetopsida сейчас представлены тремя вроде бы совершенно разными растениями: южноафриканская вельвичия Welwitschia mirabilis вообще не похожа ни на одно растение, а эфедра Ephedra подобна саксаулу с красными «ягодами-малинками», и лишь гнётум Gnetum выглядит как приличное дерево с гладкими миндалевидными листьями и красными «ягодами».
Хвойные Pinopsida – это всем известные сосны, ёлки, лиственницы, пихты и кедры, но также и некоторые растения с весьма крупными листьями, например Agathis, Nageia и Podocarpus.
Голосеменные стройными рядами начали захват планеты, что им, собственно, и удалось, учитывая, что и поныне они образуют самые огромные по площади леса.
Некоторые примитивные голосеменные ещё были внешне очень похожи на древовидные папоротники. Так, у Adiantites, Rhodeopteridium и многих других представителей класса «семенных папоротников» Lyginopteridopsida (они же Lyginopteridophyta, Pteridospermae, или Pteridospermophyta) ветки-листья так и называются вайями, а у их самых продвинутых представителей Glossopteridales – Glossopteris и Gangamopteris – были уже нормальные листья с черешком. Глоссоптериевые кустарники и невысокие деревья распространились с середины карбона, после усиления засушливости и уменьшения разнообразия плаунов и хвощей.
Самые типичные и распространённые голосеменные карбона – примитивные хвойные Pinopsida – Cordaitopsida (или Cordaitales), предки современных ёлок. Среди них особенно знаменит Cordaites, леса из которого шумели буквально по всей планете. Эти высокие стройные деревья были украшены длинными пальцевидными листьями, расходящимися от концов молодых веток характерными веерами. Показательно, что у суровых якутских кордаитов листочки не превышали 3–4 см, тогда как у расслабленных тропических еврамерийских могли вырастать до метра. Пыльцевые зёрна – микроспоры – имели один воздушный мешок. Шишек у кордаитов ещё не было, пыльца и семена образовывались в характерных чешуйчатых «почках», собранных в «серёжки».
Более продвинутые и совсем настоящие хвойные – Walchia (они же Lebachia) – кустарники и невысокие деревья Северного полушария. Их узкие вытянутые шишки ещё очень напоминали кордаитовые «серёжки», в которых отдельные «почки» были прикрыты плотно прижатыми чешуями. Листья лебахий и вальший уже превратились в знакомые нам по ёлкам и араукариям кожистые иголки. Эти растения иногда включаются в группу Voltziales, чьё торжество было ещё впереди.
С середины карбона стали набирать обороты саговниковые, или цикадовые Cycadopsida, например Spermopteris; впрочем, время их триумфа тоже ещё не настало.
В конце карбона появился Dichophyllum, в длинных узких глубоко рассечённых листьях которого только ботаник может углядеть характерные черты знаменитого потомка – гинкго.
Бурный рост крупных наземных растений, как обычно, привёл к экологическому кризису. Корни у многих тогдашних растений были ещё слабые, эта недоделка привела к сумасшедшему лесоповалу. А меж тем сосудистые растения наловчились производить лигнин – набор полимеров, цементирующий сосуды и укрепляющий крупные стебли. Только вот бактерии и грибы ещё не очень научились разлагать этот лигнин обратно. Из-за этого падающие ветки и стволы не успевали сгнивать, огромное количество древесины просто тупо накапливалось, наслаивалось само на себя, образуя невероятные толщи угля – до четверти всего угля на планете! Собственно, совсем неспроста период получил своё название – карбон, каменноугольный. Накопилось столько угля, что вот уж столетия, как люди жгут и жгут его в промышленных масштабах, а он всё не кончается. Понятно, что такой мощный вынос углерода из атмосферы не мог не сказаться на её составе: меньше углерода – больше кислорода.
Как вы там, потомки?
Разлагающийся лигнин очень характерно пахнет – это тот самый всем знакомый запах старых книг, которого так не хватает современным пользователям электроники.
А где больше кислорода, там и дышится легче, там даже на примитивном трахейном дыхании, основанном на простой диффузии, безо всяких насосов, можно надышаться до вполне приличных размеров. Не странно поэтому, что визитная карточка карбона – гигантские членистоногие.
* * *Сухопутные лёгочные скорпионы практически не поменялись. Огромными размерами – до 70 см – отличался шотландский поздневизейский Pulmonoscorpius kirktonensis.
Позднекарбоновые пауки Idmonarachne brasieri и Aphantomartus из Франции ещё не были ядовитыми и не умели плести паутину, так как не имели прядильного аппарата. Впрочем, какую-то паутину они, вероятно, таки выделяли, просто использовали её для затыкания нор и укутывания яиц. Панцирные пауки Trigonotarbida, например, Eophrynus prestvici, в карбоне вымерли, зато возникли близкие к ним рицинулеи Ricinulei, например Polyochera и Curculioides, потомки которых живут в Африке и Южной Америке в наши дни. Также в карбоне появились более-менее родственные пауки-сенокосцы Opiliones – Nemastomoides и Eotrogulus, внешне неотличимые от современных косиножек.
Как вы там, потомки?
Современные сенокосцы – кругленькие создания с очень тонкими и длинными ногами. Их брюшко заметно членистое и не отделено от головогруди стебельком, а часто брюшко и головогрудь вообще сливаются и прикрыты единым панцирем, что явно примитивно. Зато у сенокосцев трахейная система – одна из самых продвинутых среди паукообразных. Хелицеры сенокосцев оканчиваются клешнями, почти как у раков; многие сенокосцы такими клешнями выковыривают моллюсков из их раковин или даже вскрывают раковины, как консервы. Длинные тонкие ноги сенокосцев оканчиваются лапками, в которых может быть больше ста члеников. Сгибается такая нога одной мышцей с длинным сухожилием, тянущимся по всей длине до кончика лапки, а распрямляется гидравлически – под давлением гемолимфы. Поэтому мёртвый паучок всегда скукожен. Ноги держатся на честном слове, так что, если попытаться схватить сенокосца, он легко расстаётся с конечностями и убегает на оставшихся. В отличие от большинства других паукообразных, сенокосцы умеют жевать – специальными отростками на педипальпах и передних ногах. Также необычно отличие маленьких детишек сенокосцев от взрослых, развитие идёт с невыраженным, но всё же метаморфозом. Сенокосцы не ядовиты, зато имеют пахучие железы, выделяющие вонючую жидкость, которая к тому же имеет бактерицидные свойства.
Самая классная особенность сенокосцев – свойство собираться в большие тусовки, иногда из сотен особей. Когда кто-нибудь находит такой мохнатый шар из ножек в траве или углу сарая, то конечно же начинает тыкать туда палкой, отчего сенокосцы пушистой лавой, спотыкаясь и путаясь в ногах, но резво разбегаются во все стороны, а слабонервные очевидцы (вернее – очевидицы) пронзительно визжат. В общем, радость гарантирована.
Из прочей наземной мелочи продолжали копошиться многоножки, например, Euphoberia tracta. Впрочем, некоторая такая мелочь переставала быть просто козявочками: многоножка Arthropleura armata достигала длины 1,5–2,3 м, причём она была ещё и очень широкой. Трудно сказать – было ли это защитным ответом на появление хищников-амфибий или же, напротив, артроплевры сами отъедались на амфибиях до своих титанических масштабов; впрочем, чаще они реконструируются как растительноядные существа. В любом случае это были самые крупные наземные членистоногие всех времён.
Великое событие карбона – бурная эволюция насекомых Insecta. Первичнобескрылые французские Dasyleptus lucasi раньше даже выделялись в свой отряд Monura, но в наибольшей степени похожи на щетинохвосток Archaeognatha, а Carbotriplura kukalovae из Чехии – на чешуйниц Zygentoma. Показательно, что и щетинохвостки, и чешуйницы живут и размножаются только во влажных местах – они до сих пор не окончательно покинули воду. Современные первичнобескрылые насекомые – мелкие создания, а карбоновые могли достигать внушительных размеров: карботриплура вырастала до 10 см, гигантским был и Ramsdelepidion shusteri из Иллинойса.
Такие примитивные существа очень быстро обрёли крылья и стали крылатыми насекомыми Pterygota. Крылья, судя по всему, возникли лишь один раз, хотя в последующем развивались массой разных путей.
Маленькая тонкость
Пути и причины возникновения крыльев насекомых – предмет длинной и бурной дискуссии. Согласно самому ходовому предположению, крылья образовались из исходно неподвижных параноталий – плоскостей, растопыренных по бокам грудных сегментов. Такие плоскости могли использоваться как планер при прыжках с ветки на ветку. Когда крылышки подросли, насекомые научились складывать их за спиной, чтобы не слишком мешали, а отсюда уже возникла и способность активно махать при полёте. Показательно, что у многих примитивных карбоновых крылатых насекомых, кроме обычных двух пар крыльев на средне- и заднегруди, есть неподвижные крылоподобные выросты на переднегруди. Показательно, что у бескрылых чешуйниц такие же выросты используются именно для стабилизации при падениях.
По другой версии, зачатками крыльев были жаброподобные выросты на основаниях ножек. В пользу этой версии говорит строение самих крыльев – в них заходят трахеи, а также работа похожих генов на основаниях ножек ракообразных и крыльев насекомых.
Ещё одна гипотеза – крылья возникли как результат полового отбора. Возможно, самцы бескрылых насекомых трясли перед самками всякими чешуйками, половой отбор, как он это обычно делает, увеличивал эти эффектные украшалки до безумных размеров. С некоторого момента чешуйки стали знатно парусить – тут уж до крыльев было совсем недалеко. Минус этой версии в том, что крылья есть и у самок.
С появлением крыльев мир преобразился. Теперь не надо было часами забираться на высоченные деревья, можно было непринуждённо вспорхнуть на них. Расселение на огромные расстояния превратилось из подвига в банальность. Охота и спасение от хищников вышли на совершенно новый уровень.
Свежевозникшая группа древнекрылых Palaeoptera (или Odonatoptera) расцвела огромным количеством новых отрядов. Над болотами запорхали подёнки Ephemeroptera (или Ephemerida), например Triplosoba pulchella, настолько, впрочем, примитивная, что иногда выделяется в свой отряд Syntonopterida. Пыльцой кордаитов лакомились Palaeodictyoptera (или Dictyoneurida). В эту группу входит древнейшее крылатое насекомое Delitzschala bitterfeldensis с границы нижнего и среднего карбона Германии; на отпечатках его крыльев сохранились следы пёстрой окраски. Типичная палеодиктиоптера Stenodictya lobata похожа на толстую стрекозу с широкими крыльями, крылоподобными овальными выростами на переднегруди и двумя длиннющими «хвостиками»-церками сзади. Вслед за растительноядными палеодиктиоптерами понеслись стрекозоподобные Geroptera – Eugeropteron lunatum и Geropteron arcuatum, а также гигантские хищные протострекозы Protodonata (или Meganisoptera, они же Meganeuromorpha), например, древнейший чешский Erasipteron larischi и его потомки – Erasipteroides valentini и Namurotypus sippeli. Среди протострекоз особенно знаменита и замечательна позднекарбоновая Meganeura monyi – одно из крупнейших насекомых всех времён, с размахом крыльев до 65–70 см. А ведь были и многие другие – Megasecoptera (например Mischoptera nigra), Coxoplectoptera (Mickoleitia longimanus) и прочие, и прочие.
Как вы там, потомки?
Современные подёнки – удивительные насекомые, почти всю жизнь проводящие в виде яйца и личинки. Личинки живут в воде, иногда по 2–3 года. Взрослые подёнки совсем не могут питаться, даже их кишечник превратился в воздушный пузырь для облегчения парения. Понятно, что сил в таком немощном существе совсем мало, а потому взрослые подёнки живут сов-сём недолго, некоторые – один день, а некоторые так и вовсе – лишь несколько часов. Понятно, что при таком раскладе подёнки просто вынуждены вылупляться синхронно: вылет этих насекомых летним дождливым вечером над рекой представляет феерическое зрелище.
Стрекозы тоже имеют водных личинок, знаменитых своими челюстями. Нижняя губа личинки стрекозы образует так называемую маску – здоровенную выдвигающуюся членистую шкворню, которой злобная тварь может убить даже маленькую рыбку. Стрекозы – отличный пример неравномерности эволюции. С одной стороны, их крылья технически устроены крайне примитивно, неспроста их выделяют в группу древнекрылых. С другой стороны, система управления этими крыльями совершеннее, чем у большинства или даже всех других насекомых.
Тут надо сделать маленькое отступление. Например, если разрушить грибовидные тела – особые структуры мозга – саранчи, она будет без конца летать, не в силах остановиться. Это значит, что грибовидные тела тормозят импульсы полёта, возникающие в более примитивных центрах мозга. Система логична: когда враг пытается схватить саранчу, невыгодно начинать думать, что пора бы полететь. Лучше снять запрет, а сигнал всегда наготове, так реакция становится намного быстрее. Потому так трудно поймать кузнечика. Если разрушить грибовидные тела сверчка, он будет без конца чирикать, не в силах остановиться, ведь его сверчание – это начало и конец открывания крыльев для полёта, только никогда не завершаемого; крылышки трутся о ножки, вот и получается «чирик-чирик». Если разрушить грибовидные тела жука-усача, он будет бесконечно двигаться вперёд и щёлкать челюстями, не в силах остановиться. Значит, грибовидные тела тормозят движения ног и жвал.
А вот если разрушить грибовидные тела стрекозы, она замирает на месте. Это значит, что они выполняют роль именно волевого, принимающего решения центра. Стрекоза именно хочет летать, придумывает, как ей это сделать, делает это творчески. Потому-то она идеально охотится на мух и комаров, у которых идеальная механика крыльев со стабилизаторами-жужжальцами, но хуже управление на нервном уровне.
Развитие мозга стрекоз сказывается не только на полёте. У них идеальное зрение, что очевидно, если посмотреть в их невероятные глаза-шары. У них бывает довольно хитрое территориальное поведение. Понаблюдайте летом за стрекозками на пруду или ручье: каждая взлетает со своего места и возвращается строго на свой личный листочек, а не куда попало.
В рекордные сроки возникли и новокрылые насекомые Neoptera. В числе самых древних и примитивных были Paoliida, например, среднекарбоновые Kemperala hagenensis и Holasicia rasnitsyni.
Наконец, карбон осчастливил нас столь всеми любимыми тараканами Blattodea, равно как несколькими очень близкими к ним группами. Очень примитивна Manoblatta bertrandi из среднего карбона Франции, а наиболее знаменита и богата видами Phyloblatta; вообще же, известны десятки родов. Тараканы и их родственники крайне мало поменялись с карбоновых времён, ведь образ жизни в лесной подстилке с тех пор не изменился. Например, позднекарбоновый таракан Dictyomylacris insignis выглядит точь-в-точь как нынешние южноамериканские мраморные тараканы Archimandrita tesselata. Некоторые тараканоподобные насекомые, например Adeloblatta, возможно, уже относились к тараканосверчкам Grylloblattodea (или Notoptera).
Работа над ошибками
Многие глубоко убеждены, что тараканы – невероятно живучие существа, едва ли не единственные, кто способен выжить после ядерной войны. Однако в реальности в большинстве своём тараканы – довольно привередливые создания. Даже пресловутые рыжие пруссаки Blattella germanica – вроде бы такие вездесущие и неистребимые, на самом деле довольно хрупкие и чуткие. Просто их требования к жизни идеально совпадают с нашими, вот им и нравится жить в человеческих жилищах. А в террариуме держать их сложнее, чем многих тропических тараканов. За стеклом пруссаку уже и температура не та, и влажность не ся, он чахнет и гибнет в тоске по щели за кухонным косяком.
Предпрямокрылые Protorthoptera (например, Sthenaropoda fisheri) – с виду что-то среднее между тараканом и кузнечиком – в позднем карбоне доэволюционировали до прямокрылых Orthoptera, первыми представителями которых стали Oedischia.
Наконец, в конце карбона появляются насекомые с полным превращением – отряд Palaeomanteida (или Miomoptera) – германская Archaemioptera carbonaria. Внешне эти великие предки, как несложно догадаться, выглядели очень блекло, без наворотов: четыре самых банальных крылышка, незамысловатое тельце, грызущие челюсти, простенькие тоненькие усики. Яйца они откладывали в ткани растений, личинки развивались в шишках голосеменных, а взрослые питались пыльцой тех же деревьев.
Из Мазон Крик в Иллинойсе с древностью 308 млн л. н. происходят отпечатки вроде бы самого древнего известного жука – Adiphlebia lacoana. Впрочем, некоторые исследователи считают его слишком особенным и, хотя признают, что он относится к насекомым с полным превращением, но исключают из жуков.
С этого момента планета была обречена на захват насекомыми.
Маленькая тонкость
Современные насекомые иногда упрощённо делятся на две большие группы: насекомых с неполным превращением Hemimetabola и насекомых с полным превращением Holometabola. Необходимо отметить, что в большинстве современных систематик такое деление не принимается, так как группа с неполным превращением явно сборная. Однако в первом приближении такая схема удобна, отчего и принимается, например, в школьном курсе зоологии.
У насекомых с неполным превращением личинка (по-умному – нимфа) много раз линяет, с каждой линькой чуток подрастая, наращивая крылья и потихоньку превращаясь во взрослую форму (по-учёному – имаго). У насекомых с полным превращением личинка очень сильно отличается от взрослой формы, причём не только внешне, но и образом жизни, в том числе типом питания. Это позволяет избежать конкуренции нимф и имаго и на одной территории существовать вдвое большему числу особей. В некоторый момент личинка окукливается, внутри куколки все её ткани фактически растворяются, а потом заново пересобираются в совершенно новый организм взрослого насекомого.
Насколько это известно, настоящее полное превращение возникло единожды, хотя попыток было много, можно вспомнить хотя бы подёнок, стрекоз или белокрылок, у которых, впрочем, нет куколки.
* * *Если есть столько членистоногой еды, то неизбежно появятся её потребители. Стрекозоподобные твари не справлялись, им на помощь устремились амфибии. Как и насекомые, амфибии в рекордные сроки дали великое разнообразие. Разобраться в нём нелегко. Раньше всё было просто – всех каменноугольных амфибий скопом называли «стегоцефалами», теперь же выделяется около десятка линий, крайне туманно связанных между собой. Есть даже мысли, что разные амфибии независимо возникли из разных рипидистий или двоякодышащих, хотя и для этой точки зрения чётких доказательств нет. Важнейший для систематики амфибий признак – строение позвонков. У многих примитивных амфибий эмаль зубов была изборождена очень сложными бороздками, как у рипидистий, отчего в упрощённом варианте классификации их объединяют в одну группу Labyrinthodontia.
Маленькая тонкость
Строение позвонков амфибий – одна из самых значимых для классификации особенностей. Известна куча вариантов.
Рахитомный тип – самый примитивный, доставшийся ещё от рипидистий: тело позвонка образовано передне-нижним непарным серповидным гипоцентром под хордой и задним парным плевроцентром сверху и по бокам от хорды; невральные дуги самостоятельны.
Стереоспондильный тип: тело образовано дисковидным гипоцентром, плевроцентры исчезают; невральные дуги соединены с телом швами.
Эмболомерный тип: тело состоит из двух дисков, один – из гипоцентра, другой – из плевроцентров; невральная дуга вклинивается между ними.
Гастроцентральный (или плевроцентральный) тип: плевроцентральный диск увеличен, а гипоцентральный уменьшается до серповидного интерцентра внизу, без связи с невральной дугой.
Апсидоспондильный (или нотоцентральный) тип: тело позвонка сформировано разросшимся основанием невральной дуги, а плевроцентров и гипоцентра вообще нет.
Лепоспондильный тип: тело развивается в виде кольца поверх хряща вокруг хорды.
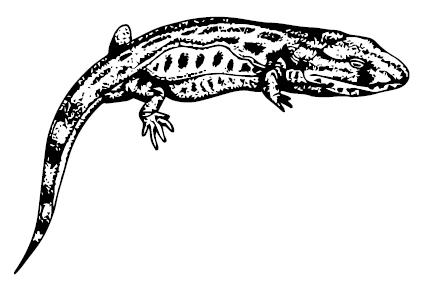
Caerorhachis bairdi
Temnospondyli – одна из самых богатых групп лабиринтодонтов. Продвинутой чертой темноспондилов было развитие двух шейных позвонков (у современных амфибий он один). К темноспондилам, в частности, близки Colosteus: у этих примитивных амфибий череп был похож на ихтиостежий, тело удлинено, лапки уменьшены, а на брюхе сохранялась чешуя; сходно выглядели Ossinodus pueri и Greererpeton buckemorani. Dendrerpeton и Caerorhachis bairdi были подобны тритонам, Loxomma имела широкое рыло и огромные глаза. В середине карбона появился «лягушкозавр» Stegops – первый представитель Eryopoidea (или Eryopsoidea), группы, столь значимой в пермском периоде. Какие-то темноспондилы, вероятно, стали предками современных бесхвостых амфибий. Несколько особняком стоят лабиринтодонты Baphetidae, например Megalocephalus, предполагавшиеся в качестве предков рептилий, хотя против этого есть свои соображения.
Anthracosauria – другая важнейшая группа лабиринтодонтов, она включает в себя Embolomeri. Образцовый пример – Proterogyrinus – толстое существо с вытянутым приплюснутым черепом и довольно крупными лапами. Некоторые эмболомеры повторили судьбу темноспондилов: у Pholidogaster pisciformis и Eogyrinus attheyi тело вытянулось, лапки укоротились, а живот был покрыт костной чешуёй. Среднекарбоновый шотландский Pteroplax cornuta имел голову длиной 40 см, а сам вырастал до 4–5 м – одно из самых больших животных палеозоя. Скорее всего, именно какие-то антракозавры стали предками рептилий, по крайней мере, лучших кандидатов нет.
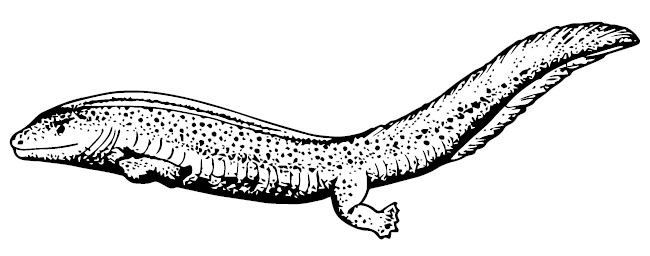
Eogyrinus attheyi
Lepospondyli, наверное, самые странные стегоцефалы. Их позвонки формировались в виде костного кольца вокруг хорды, а зубы потеряли складчатость. Если в группе Nectridea примитивные Scincosaurus и Ptyonius мало отличаются от тритонов, то особо зубастый Urocordylus имел выступы на задних углах черепа, а продвинутые треугольноголовые Batrachiderpeton отрастили из таких выступов шипы-рога, несколько расходящиеся в стороны, тогда как у трапециеголовых Diceratosaurus подобные рога торчали строго назад. Рога совершенно недвусмысленно указывают на появление хищников. Потомки нектридей в перми довели рогатую идею до крайности.
Другая группа лепоспондил – Aistopoda: Dolichosoma и Lethiscus stocki были червеподобными крайне тонкими животными со 100–250 позвонками и совсем без ножек, покрытыми сверху и снизу чешуйным панцирем. Такими же длинными и безногими (плечевой пояс при этом сохранялся) и с мелкими костными чешуйками на животе были Adelospondyli – Palaeomolgophis scoticus и Adelospondylus watsoni. Близкие к ним Lysorophia, например, Lysorophus, Cocytinus и Brachydectes, всю жизнь сохраняли окостеневшие жаберные дуги. Все эти вытянутые создания копошились в придонном иле и процеживали воду между своими мельчайшими зубками.
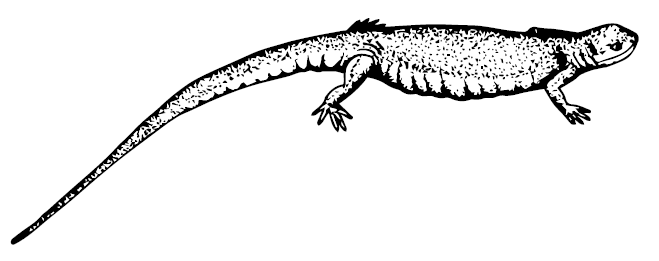
Scincosaurus
Ещё одна группа – Microsauria; их череп и зубы похожи на таковые у лепоспондилов, но позвонки устроены несколько иначе, а тело покрыто чешуйным панцирем. Большинство микрозавров стали неотеническими формами, всю жизнь дышавшими наружными жабрами, подобно современному аксолотлю. Отличные примеры – Microbrachis, Tuditanus, Hyloplesion и Adelogyrinus simоrhуnchus.
Некоторые карбоновые амфибии вообще не родственны никому. И виной этой неопределённости не всегда недостаток данных. Например, визейский Crassigyrinus scoticus – короткоголовый пучеглазый полутораметровый стегоцефал с крошечными лапками – известен по более-менее полному скелету, но всё равно никто не знает, к кому его отнести. По некоторым признакам крассигиринус оказывается едва ли не примитивнее вентастеги, акантостеги и ихтиостеги, так что его иногда выделяют в особый отряд Palaeostegalia.