
Палеонтология антрополога. Три эры под одной обложкой
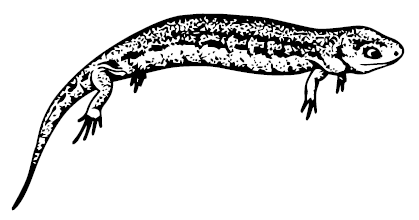
Hyloplesion
Ещё более загадочен Hesperoherpeton garnettense из Канзаса, иногда выделяемый в собственный отряд Plesiopoda. Несмотря на позднекарбоновый возраст, он имел череп из двух блоков с суставом, хорду, заходящую в череп, и боковую линию в каналах черепа – всё как у рипидистий. Вероятно, у него ещё не было среднего уха, зато имелся рудимент жаберной крышки и внутренних жабр. Передняя лапа крайне специфична: запястье состояло из одного ряда костей, каждый из четырёх пальцев нёс всего по две фаланги. На концах длинных костей передней лапы имелись характерные для рипидистий выросты, так что рука гесперогерпетона оказывается примитивнее, чем у ихтиостеги!
Как вы там, потомки?
Современные амфибии, или земноводные Amphibia делятся на три отряда: хвостатые Caudata (или Urodela; тритоны и саламандры), безногие Gymnophiona (или Apoda; червяги), а также бесхвостые Anura (или Salientia; лягушки, жабы и квакши; вместе с более примитивными вымершими группами они составляют Salientia).
Все особенности амфибий связаны так или иначе с передвижением и дыханием. Большинство современных амфибий, кроме червяг, полностью утратили чешую, ведь при несовершенстве почти гладких лёгких хорошо бы дышать кожей. Фишка амфибий – изобилие многоклеточных кожных желёз, в том числе ядовитых; особенно знамениты на этом поприще южноамериканские древесные квакши-листолазы Phyllobates.
Оттого что хоаны у амфибий расположены спереди и твёрдого вторичного нёба нет, глазницы снизу открываются прямо в ротоглоточную полость, так что лягушки используют глаза для глотания.
Из-за сочетания лёгочного и кожного дыхания кровеносная система амфибий, как ни странно, деградировала. У рыб венозная кровь идёт из сердца в жабры, насыщается кислородом, становится артериальной и прямиком следует в мозг. У амфибий же кровь из сердца идёт в лёгкие, а потом опять в сердце, так как мощности сердца не хватит, чтобы одним толчком гнать кровь до лёгких и сразу из них дальше по всему организму. В итоге артериальная кровь в сердце смешивается с венозной, которая попадает ровно туда же, только через другое предсердие. Казалось бы, надо просто разделить два круга кровообращения, как это и сделано у птиц и млекопитающих, но тут-то и мешается кожное дыхание, ведь кожа тоже насыщает кровь кислородом, но включена в большой круг кровообращения. Поэтому у амфибий кровь в желудочке оказывается смешанной, и мозги получают не самую артериальную кровь, то есть система устроена хуже, чем у рыб! Но в карбоне на суше всё равно не было никого круче амфибий, так что – «и так сойдёт!» Вот когда появились рептилии, у которых возникла хотя бы неполная перегородка желудочка (на самом деле, у амфибии карликового сирена Siren intermedia она тоже возникла, но, очевидно, намного позже), то амфибии оказались в явном проигрыше и катастрофически сократились в числе и разнообразии. Но пока – в карбоне – всё было отлично.
Как уже говорилось, в серпуховском веке случилось мощное вымирание, основательно прошедшееся и по амфибиям. Но свята экологическая ниша пуста не бывает – на место исчезнувших стегоцефалов устремились потомки выживших, закалённые в холоде ледникового периода первые рептилии – провозвестники нового миропорядка.
А где же люди? Первые рептилии
Уже с начала карбона разные амфибии начали пытаться превратиться в рептилий. Огромные леса заполонили огромные пространства, там бегали мириады вкусных тараканов, всё это очень хотелось съесть, но была большая проблема: весь организм амфибий, а особенно икра и личинки-головастики, был зависим от воды. Растения и насекомые намного раньше решили подобные проблемы и подготовили среду для окончательного выхода четвероногих на сушу. Наставало время рептилий.
Проблемы с сохнущей икрой были решены развитием и совершенствованием зародышевых оболочек, окруживших уже не икру, а яйцо.
Хорион – самая наружная оболочка, стала затвердевать и превратилась в скорлупу, известковую или кожистую. Это, очевидно, механическая защита.
Маленькая тонкость
Скорлупа только кажется такой уж непроницаемой. На самом деле она пронизана множеством маленьких пор. На этом основан способ сохранения прохладной воды в пустыне: берётся страусиное яйцо, содержимое, понятно, съедается, а в скорлупу через дырочку заливается вода. Скорлупа через микропоры помаленьку пропускает отдельные «самые горячие» молекулы, которые испаряются с поверхности, унося энергию, а вода внутри остаётся прохладной.
Амнион – тонкая плёночка под скорлупой, как раз почти водонепроницаемая, служит для защиты от высыхания. Амнион окружает амниотическую полость, заполненную амниотической жидкостью, то есть получается свой маленький личный аквариум, в котором плавает довольный зародыш. Разница лягушки и ящерицы не в том, что у первой есть стадия головастика с жабрами, а у второй – нет. Разница сугубо в сроках и последовательности событий. У лягушки сначала из икринки вылупляется головастик с жабрами, одним предсердием и боковой линией, без ног и скелета – фактически круглоротая рыба, а потом происходит метаморфоз и все переделывается на взрослый лад. У ящерицы же (да и у нас тоже) сначала эмбрион проходит метаморфоз – исчезают зачаточные жабры и боковая линия, появляются второе предсердие, скелет и ручки-ножки, а потом уже готовый детёныш вылупляется из яйца. У лягушки головастик плавает в водоёме, и у нас тоже, только у нас этот водоём ограничен амнионом.
Существа, имеющие амнион – амниоты, это рептилии, птицы и млекопитающие; в противоположность им, рыбы и амфибии – анамнии. В этом смысле лягушка ближе к акуле и карасю, чем к ящерице и человеку.
Аллантоис служит главным образом для дыхания зародыша, но на ранних стадиях развития важен и как запасающий воду пузырь. Более того, он способен втягивать воду из окружающей среды, так что у некоторых ящериц и черепах с момента откладки яйца до вылупления детёныша количество воды в яйце увеличивается вдвое.
Как вы там, потомки?
Некоторые современные безлёгочные саламандры семейства Plethodontidae, например, родов Desmognathus, Plethodon, Hemidactylium и Batrachoseps, демонстрируют, как могли возникнуть яйца: они откладывают крупные икринки прямо на землю во влажных местах и обвивают их своим телом. Вылупившиеся личинки, хотя и дышат жабрами, не живут в воде, а ползают по мокрым кочкам и быстро проходят метаморфоз.
Антильская листовая лягушка Eleutherodactylus martinicensis пошла в некотором отношении дальше. Она откладывает икринки-яйца, заключённые в общую оболочку, в пазухи листьев. Всё развитие детёныша происходит внутри икринки без метаморфоза, у личинки нет наружных жабр, а дыхание происходит через капилляры укороченного и расширенного хвоста, прилегающего к оболочке икринки.
Даже у некоторых костистых рыб возникает подобие яиц: африканские Nothobranchius и Aphyosemion откладывают икру в сравнительно сухих местах, а некоторые даже в дуплах деревьев, причём без высыхания икра в принципе не развивается. Оболочка-хорион у таких икринок утолщена и не позволяет сохнуть, так что стала аналогом амниона.
Кроме оболочек, яйцо усовершенствовалось увеличением количества желтка, ведь зародыш должен развиваться внутри скорлупы до вполне приличного состояния, да к тому же не особенно стучаться об эту скорлупу, так что желток выполняет не только питательную, но и амортизационную роль.
Благодаря всем этим наворотам четвероногие перестали зависеть в своём размножении от воды.
Маленькая тонкость
Как обычно, у всего есть и обратная сторона. Безопасность размножения рептилий сделала ненужной заботу о потомстве. У рыб и амфибий её, конечно, обычно тоже нет, но всё же примеров достаточно. А у рептилий с заботливостью всё совсем плохо: у большинства видов яйца даже не закапываются в землю, а просто откладываются куда попало на землю или листья – «и так сойдёт». Минимальную ответственность изображают лишь крокодилы, питоны и королевские кобры. У крокодилов это выражается в том, что крокодилица закапывает яйца в кучу песка и листьев (в книгах это называется гнездом, но в реальности всё выглядит не так красиво), ложится сверху и лежит (в книгах сказано, что она охраняет кладку, но в действительности понятно, что просто дураков лезть под злобную зубастую тварь нет, хотя мангусты умудряются воровать яйца даже при таком раскладе). В книгах обычно ещё пишут, что мамаши тонко регулируют температуру в гнезде, но стоит посмотреть на оригинал: крокодилица иногда лениво дрыгает то одной лапкой, то другой, отгребая порцию грязи. Когда крокодильчики вылупляются и самовыкапываются, мамочка крайне аккуратно берёт их в рот и выпускает в воду. С этого момента её родительский долг полностью исчерпан; если на следующий день крокодилиха случайно встретит детёныша, то без затей сожрёт его – отбор, однако.
Питонихи обвиваются вокруг кладки и даже, будучи холоднокровными, но мускулистыми, могут нагревать яйца на 2–4 °C. Королевские кобры Ophiophagus hannah тоже охраняют гнездо – кучу листьев, но заключается охрана просто в присутствии ядовитой гадины около яиц.
Чтобы не зависеть от воды и во взрослом состоянии, кожа рептилий стала сухой, без желёз, ороговевшей в виде рептильной чешуи, которая совсем не то же самое, что чешуя рыбья. Кожное дыхание свелось к минимуму, зато воздух в лёгкие стал не закачиваться движениями дна ротовой полости, а всасываться расширением грудной клетки. Жизнь на суше поменяла всю форму тела: голова уменьшилась и сузилась (площадь ротового дна уже не важна, а укус узкими высокими челюстями получается сильнее), шея и, соответственно, трахея с пищеводом удлинились, туловище укоротилось, позвоночник стал прочнее за счёт полного окостенения, а хорда большей частью пропала (хотя сохраняется у гаттерии и гекконов), живот укрепился брюшными рёбрами (образовавшимися из окостенений кожи на рыбьем пузике), а таз стал крепиться не к одному, а к двум крестцовым позвонкам, лапы вытянулись и окрепли, пальцы удлинились, а на их кончиках выросли когти. В силу возросшей бодрости менялась координация движений и работа мозга, а хрусталик глаза стал, наконец, эластичным. Судя по современным рептилиям, в желудочке сердца появилась неполная перегородка, а обмен веществ перестроился на водоэкономящее выделение мочевой кислоты, а не мочевины.
Все эти процессы шли не одновременно и в разных группах с разными скоростями, попыток «рептилизоваться» было много. Одна из первых принадлежала раннекарбоновым Whatcheeriidae – турнейскому шотландскому Pederpes finneyae и визейской айовской Whatcheeria deltae – метровым крупноголовым пучеглазым саламандроподобным существам с довольно большими лапами, на которых они могли довольно бодро бегать по земле. Сравнительно узкий череп педерпеса был плохо приспособлен для гулярной ундуляции, то есть воздух в лёгкие он накачивал не горлом, а уже рёбрами. Таким образом, педерпес вполне годится на роль Великого Предка всех наземных четвероногих. Вместе с тем, на его кисти сохранялся тоненький предпервый палец – привет от тулерпетона.
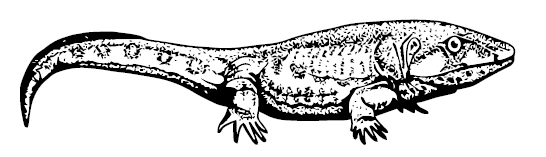
Pederpes finneyae
В визейском веке раннего карбона в Шотландии появляются первые «почти-почти совсем рептилии» – Casineria kiddi, Westlothiana lizziae, Eldeceeon rolfei и Silvanerpeton miripedis. Как положено Великим Предкам, это были весьма скромные животные длиной 15–40 см, самого невзрачного вида, очень похожие на ящериц, со сравнительно укороченным туловищем, длинной для амфибий шеей (четыре шейных позвонка у вестлотианы), двумя крестцовыми позвонками и увеличенными пятипалыми ногами (только у вестлотианы туловище длинное, а ножки короткие), а в целом – с мозаикой амфибийных и рептилийных черт. Существенно, что отложения, в которых найдена казинерия, сформировались в сухих условиях.
В середине карбона появляются первые совершенно несомненные рептилии Captorhinida (или Captorhinomorpha). Без сомнения, главный Великий Предок тут – Hylonomus lyelli из московского века (314 млн л. н.) Канады. Эта ящерицеподобная зверюшка длиной 20–25 см, судя по множеству мелких зубок, питалась насекомыми.
Маленькая тонкость
Скелет гилономуса обнаружен внутри пня древовидного плауна Sigillaria. Видимо, во время наводнений мелкие существа попадали в такие трубы-ловушки, а после спада воды не могли вскарабкаться по гладким стенкам. Зато палеонтологам достались окаменелости идеальной сохранности. Кроме костей, найдены и следы пятипалых лапок наземных «ящериц», которые, скорее всего, принадлежат тем же самым гилономусам.
Едва ли не примитивнее гилономуса, но на несколько миллионов лет позже его по времени канадский Paleothyris acadiana и чешская Brouffia orientalis. В целом же они выглядели примерно так же – и по размерам, и по внешнему виду, и по образу жизни. Следующую ступень преодолели Cephalerpeton ventriarmatum из касимовского века Иллинойса и Anthracodromeus longipes из Огайо. Coelostegus prothales из касимовского века Чехии даже несколько примитивнее предыдущих рептилий; для звания Великого Предка ему опять же не хватает древности. Линию продолжает Euconcordia cunninghami (она же Concordia, хотя это название раньше было дано одной креветке) из гжельского века Канады; правда, её потомки-капториниды после зашли в эволюционный тупик.
Наконец, уже в середине и конце карбона обнаруживаются очередные Великие Предки всех главных групп последующих рептилий.
Парарептилии Parareptilia, или анапсиды Anapsida, от которых, в частности, произошли черепахи. Одна из их важнейших особенностей – отсутствие височных окон в черепе.
Зауропсиды Sauropsida, они же настоящие пресмыкающиеся Eureptilia, или диапсиды Diapsida – большинство рептилий, от которых, в частности, позже произошли птицы. Их особенность – две пары височных окон в черепе.
Синапсиды Synapsida, или зверообразные Theromorpha – предки млекопитающих. У них в черепе пара височных окон – по одному с каждой стороны.
Маленькая тонкость
Височные окна – отверстия в черепе между глазницей и ухом – возникали не один раз, независимыми путями и между разными костями. Их смысл – разгрузка напряжений, возникающих из-за жевательной мускулатуры. Между прочим, и ушные вырезки на заднем крае черепа для прикрепления барабанной перепонки тоже эволюционировали массой способов, их особенностям посвящены сотни страниц специальной литературы.
Первые представители парарептилий – Desmatodon hollandi и D. hesperis из касимовского века США, относимые к Diadectomorpha и даже точнее – Diadectidae; в синхронных слоях Германии найдены следы лап подобных животных, названные Ichniotherium praesidentis. Они же по совместительству были первыми растительноядными четвероногими. Таким образом, с этого момента можно начинать отсчитывать новую эпоху – потребления растений не только грибами и беспозвоночными, но и наземными позвоночными.
Древнейшими зауропсидами были Spinoaequalis schultzei и Petrolacosaurus kansensis из гжельского века (302 млн л. н.) Канзаса, включаемые в отряд Araeoscelidia. Внешне они выглядели как сорокасантиметровые ящерицы с длинными стройными пятипалыми лапками, брюшными рёбрами и двумя парами височных окон, отчего уверенно определяются как предки всех диапсид. Кроме прочего, первые рептилии заметно отличаются от амфибий большим числом шейных позвонков, например, у петролакозавра их было шесть или семь. К сожалению, далеко не про всех ископаемых животных известны такие подробности. Судя по зубам, петролакозавр был насекомоядным, а судя по длине и форме пальцев, мог лазать по деревьям. Спиноэквалис по части признаков даже примитивнее, но, видимо, снова научился неплохо плавать, гребя очень длинным плоским хвостом.
В одном местонахождении с гилономусом найдены фрагментарные остатки пеликозавра Protoclepsydrops haplous и другой зверообразной рептилии Novascoticus multidens (впрочем, новаскотикус мог быть и амфибиеймикрозавром). Выходит, что звероящеры появились на самой ранней заре пресмыкающихся, уже 314 млн л. н. Не странно, что они сохранили часть признаков амфибий. И это очень хорошо, так как стало залогом многих ключевых особенностей млекопитающих, включая, собственно, млекопитание.
В той же Канаде 308 млн л. н. жил Echinerpeton intermedium, а 306 млн л. н. – Великий Предок Archaeothyris florensis. Если систематическое положение первого из-за плохой сохранности остатков ещё довольно спорно, второй уверенно определяется как член семейства пеликозавров Ophiacodontidae. Среди острых зубов археотириса выделялись размером «клыки» – первая ласточка нашей гетеродонтности. Форма черепа археотириса уже узнаваемо зверообразная: вытянутая узкая морда, большие глазницы, по одному височному окну с каждой стороны, опущенный относительно зубного ряда затылок – для более широкого разевания пасти. Очевидно, археотирис был самым злым хищником лепидодендровых болот – наше исходное призвание.
Возможно, даже самые первые пеликозавры уже иногда могли заботиться о потомстве, по крайней мере, на это намекает находка в московском ярусе Канады ассоциированных скелетов взрослого и детёныша Dendromaia unamakiensis из семейства Varanopsidae (или Varanopidae). Внешне эти ящерицеподобные существа были невзрачны, но даже самые заурядные существа могут порой удивить.
Известны и следы ног зверообразных пеликозавров из касимовского века Германии, названные Dimetropus. По ним отлично видно, что эти животные передвигались ещё не очень быстро на широко расставленных лапах, но их пальцы уже были направлены строго вперёд.
А в самом конце карбона, в гжельском веке Канзаса, уже существовали несколько видов Ianthasaurus – мелких насекомоядных пеликозавров, родственников эдафозавров, с высоким гребнем на спине из удлинённых остистых отростков позвонков.
* * *Несмотря на появление настоящих рептилий, в конце карбона часть групп амфибий продолжала эволюционировать в том же направлении. Gephyrostegidae (иногда включаемые в антракозавров, а иногда выделяемые в свой отряд Gephyrostegida), например Bruktererpeton fiebigi, Eusauropleura digitata, Gephyrostegus bohemicus и G. watsoni, имели узкий череп, довольно длинные пятипалые ноги, короткое тело с брюшными рёбрами и костными чешуйками на спине и животе. Это были мелкие насекомоядные ящерицеподобные животные, которые часть жизни вполне могли проводить в воде, но уже были почти совсем наземны. Единственная проблема гефиростегид – они опоздали, рептилии уже появились и надёжно заняли свои экологические ниши. Плохо окостеневший позвоночник и несколько тяжеловатая голова были явными минусами для наземной жизни. Практически то же можно повторить о чуть более продвинутых Solenodonsauridae, например о чешском Solenodonsaurus janenschi, и некоторых других животных.
* * *Конец карбона, как полагается, ознаменовался вымиранием. В позднем карбоне климат стал совсем грустным, наступал среднекарбоново-пермский ледниковый период, уменьшались как температуры, так и влажность. Изменения были постепенными. На границе московского и касимовского веков заметно уменьшилось количество древовидных плаунов, хвощей и кордаитов, но зато увеличилось число папоротников, в том числе древовидных. К концу касимовского века леса полностью поменяли свой облик. Тропические леса, покрывавшие до того огромные территории, большей частью вовсе исчезли, а оставшиеся распались на отдельные островки. С одной стороны, это привело к появлению изолированных эндемичных фаун, с другой – приспособленные к размножению в сухих местах рептилии получили возможность быстро расселяться на большие расстояния, благо континенты продолжали слипаться всё теснее и теснее.
Похолодание, осушение и снижение количества кислорода в атмосфере – ведь фотосинтезом с исчезновением лесов стало заниматься меньше растений – привели к сокращению числа гигантских насекомых и многоножек. Да и пищи для членистоногих стало меньше. В итоге, исчезли многие палеодиктиоптеры, а вслед за ними – масса стрекозоподобных хищников, хотя, конечно, тоже не все. Понятно, что за насекомыми на границе московского и касимовского веков последовала в могилу и большая часть их потребителей – амфибий. А вот на рептилиях эти катаклизмы не только не сказались отрицательно, напротив, число видов их со временем только росло.
Следующий – уже третий для карбона – пик вымирания пришёлся на границу с пермским периодом. Тут как раз пострадали в основном рептилии, тогда как оставшиеся амфибии едва ли не приросли в числе. Правда, наземные фауны на границе карбона и перми изменялись как раз не так выразительно, как на границе раннего и среднего, а также среднего и позднего карбона. В очередной раз становится очевидно, что проведение рубежей периодов – дело относительное.
Вымирали не только наземные флоры и фауны, во второй половине карбона почти полностью исчезли рипидистии и значительная часть акантод.
Как обычно, выдвинута и катастрофическая версия позднекарбонового вымирания, привязывающая его к извержениям в районе нынешней Скандинавии, хотя хронологически извержения случились позже вымираний.
* * *Карбон вывел нас из болот на просторы суши, превратил икру в яйца, высушил слизистую кожу, вытянул нашу шею и привёл в движение наши рёбра, дал нам наземное зрение и длинные ноги. Карбон вселил в нас недоверие к многоножкам и заложил основы насекомоядности, сыгравшие столь важную роль в эволюции млекопитающих в целом и приматов в частности. Карбон – этот период-помойка, заваливший весь мир гниющими стволами – снабдил нас безмерными запасами угля для энергетического обеспечения нашей цивилизации и тем обеспечил промышленную революцию XIX века.
АльтернативыВ карбоне расцвели амфибии и появились рептилии. Такого изобилия и разнообразия четвероногих, таких перспектив и возможностей не было ни до, ни после карбона. Вся планета лежала перед пионерами суши. Все имели равные шансы. Кто-то упустил их, распрощавшись с ногами или став неотеническими личинками, но кто-то до последнего пытался занять своё место в истории. Изобилие рептилизовавшихся карбоновых амфибий поражает, учитывая ограниченность наших знаний. Ясно, что в реальности таких эволюционных линий было намного больше. Наши предки были далеко не единственными претендентами на захват планеты. Даже среди первых зверообразных уже появились несколько альтернативных вариантов.
А могло быть и совсем по-другому. Например, позднедевонские или раннекарбоновые амфибии могли быстро распрощаться с рёбрами, как это в последующем сделали лягушки и жабы, и никогда не развить рёберного дыхания. Тогда активный обмен так и не появился бы, и до сих пор по болотам ползали бы саламандроподобные скользкие твари, лениво ловя тараканов и кузнечиков. Амфибии могли слишком быстро потерять ноги, как это случилось с аистоподами; они, конечно, тоже освоили бы сушу и рептилизовались, но Земля превратилась бы в планету змей, которые никогда не освоили бы трудовую деятельность и не стали бы разумными. Кто знает, может, тогда свой призрачный шанс реализовали бы какие-то членистоногие?
Гигантские протострекозы могли усовершенствовать свои ганглии, отрастить грибовидные тела побольше, стать нелетающими, ещё больше вырастить мозги, а там, глядишь, и до разума недалеко. Как бы выглядела цивилизация таких насекомых?
Пермь
298,9–251,902 миллиона лет назад: Зима палеозоя, взлёт и падение звероящеров
МЕЖДУНАРОДНАЯ (И РОССИЙСКАЯ) ШКАЛА:
298,9 млн л. н. – приуральская эпоха: ассельский век – 293,52 – сакмарский век – 290,1 – артинский век – 283,5 – кунгурский век (кунгурский век, уфимский век) – 272,95 – гваделупская (биармийская) эпоха: роудский (казанский) век – 268,8 – вордский (уржумский) век – 265,1 – (татарская эпоха) кептенский (северодвинский) век – 259,1 – лопинская эпоха: вучапинский (вятский) век – 254,14 – чансинский век – 251,902

Пермский период – самый холодный период палеозоя и один из самых холодных за всю историю планеты. Ужасный климат не мог не подтолкнуть эволюцию – неспроста именно в перми появились все предпосылки для возникновения млекопитающих. Ещё немного – и звери бы окончательно сформировались и заняли планету, а разум мог воссиять на сотни миллионов лет раньше. Но пермь – и самый трагический период: в его конце почти все достижения палеозоя оказались уничтожены, причём, как часто бывает с самыми ужасными событиями, этот катаклизм к тому же до сих пор малоизвестен большинству людей.