
Палеонтология антрополога. Три эры под одной обложкой
Fieldia lanceolata, Selkirkia columbia
Приапулид Priapula (или Priapulida) иногда называют головохоботными. Эти толстые морщинистые черви живут в U-образных норках, из которых высовывают длинный шипастый хобот. Особенность приапулид в том, что снаружи приапулиды выглядят как радиально-симметричные, тогда как внутреннее строение билатерально. Видимо, примитивной группой приапулид были Palaeoscolecida, например, Palaeoscolex sinensis и Cricocosmia jinningensis. Кембрийские приапулиды крайне многочисленны, но не слишком разнообразны – например, китайские Palaeopriapulites parvus, Protopriapulites haikouensis и Paraselkirkia jinningensis, якутские Vladipriapulus malakhovi, Piloscolex platum и Wronascolex lubovae, австралийская Shergoldana australiensis, канадские Ancalagon minor, Fieldia lanceolata, Selkirkia columbia, Louisella pedunculata и Ottoia prolifica, а также многие-многие другие.
Как вы там, потомки?
Современные приапулиды Priapula внешне и по образу жизни чрезвычайно похожи на совсем неродственных им сипункулид Sipuncula, эхиурид Echiura и кишечнодышащих Enteropneusta, что является отличным примером конвергенции. Все они роют норы в песке на дне или в прибойной зоне, откуда высовывают хобот и задний конец. Черви ловят пробегающих мимо животных или пропускают через себя песок, переваривают всё съедобное и выталкивают стерильный грунт наружу. Периодически какой-нибудь турист находит на пляже земляную колбасу, самопроизвольно вылезающую из песка, тут же снимает это на телефон и скорее вешает в интернете с комментариями «Что бы это значило?!». Самые отважные решаются раскопать источник удивления, с ужасом обнаруживают там огромного червяка и в панике орут дикими голосами. Успех видео гарантирован.
Тихоходки Tardigrada – уникальные существа, знаменитые способностью выживать в любых условиях.
Как вы там, потомки?
Тихоходкам нипочём 10 лет высыхания до 1 % воды от нормального, 30 лет пребывания при -20 °C, 8 часов в жидком гелии при -271 °C и некоторое время при -272 °C (это на 1 несчастный градус теплее абсолютного нуля!), 20 месяцев в жидком кислороде при -193 °C, 10 часов при +60–65 °C, 1 час при +100 °C и некоторое время при +151 °C. Они отлично чувствуют себя в атмосфере сероводорода и углекислого газа, при давлении в 6000 атмосфер, переносят облучение в 570 тысяч рентген (выживает половина тихоходок, тогда как для человека смертельны 500–600 рентген). На теневой стороне космической станции в открытом космосе упорные зверюшки продержались 10 дней. И только когда их вывесили не только в открытый космос, но и на солнечную сторону и хорошенько прожарили радиацией, почти-бессмертных существ таки удалось заморить.
Благодаря невероятной живучести тихоходки вездесущи – они живут на морском дне на глубине в 4690 м и на вершинах Альп и Гималаев выше 6000 м, в горячих источниках Японии при 40 °C и подо льдом Шпицбергена и Антарктиды. Большинство известных видов обычно заселяют капельки воды на мху – они до сих пор не знают, что вышли на сушу, ведь для них капля – тот же самый водоём.
Такие сверхспособности не возникают просто так. Жизнь в кембрии ютилась в воде, а суша представляла собой гольную пустыню – камень, песок, то много воды, то великая сушь. Наземных растений не было, почвы, соответственно, тоже, а потому земля постоянно размывалась ливнями, песок перемывался, берега непрерывно перестраивались. В этом аду, иногда раскалённом, иногда затапливаемом, могли выжить только суперсущества. И тихоходкам не оставалось ничего другого, как выработать невероятную жизнестойкость. Конечно, у всего есть и обратная сторона. Обмен веществ у тихоходок крайне низок, неспроста они носят своё название. Древнейшая тихоходка (к сожалению, безымянная) найдена в среднекембрийском местонахождении Куонамка в Якутии – и внешне она уже ничуть не отличалась от современных потомков (впрочем, есть версия, что это была личинка какой-то ксенузии). Как часто бывает в эволюции, идеальное приспособление к среде делает ненужной последующую эволюцию. Парадоксальным образом тихоходки, освоившие жизнь в максимальной нестабильности условий, нашли в этом свою стабильность. Впрочем, есть альтернативный взгляд, согласно которому именно какие-то тихоходки стали предками членистоногих, а ныне живущие представляют просто застрявших в развитии живых ископаемых.
* * *Отдельная странная группа тварей иногда объединяется в класс Dinocaridida, отряд Radiodonta и даже общий подотряд Anomalocarida, хотя и один отряд на всех кажется слишком тесным, не говоря о подотряде.
Наиболее примитивно была устроена раннекембрийская Kerygmachela kierkegaardi из Сириус Пассет. Её тело характерно для динокаридид: плоское, с лопастями по бокам, не очевидно подразделённое рядами бугорков и поперечных морщинок, так что трудно сказать, сегментировано оно или нет; снизу у неё торчали коротенькие конические парные «ножки» (впрочем, их назначение неведомо). Была, конечно, и специфика: вместо хвостовых лопастей, типичных для более поздних представителей группы, тянулся длиннющий несегментированный хвост-ость (изначально реконструированный как два длиннющих сегментированных хвоста-церки), а спереди по бокам не отделённой от тела головы растопыривались две огромные толстые «руки» с шипами, на конце становящимися длинными антеннами. Керигмачела имела вытянутые по сторонам головы глаза, собранные из множества простых глазков, но ещё не вполне фасеточные. Такая тварь достигала в длину 17,5 см и была, видимо, хищником.
Особо безумно выглядела Opabinia regalis – шестисантиметровая сегментированная «креветка» с широкими ребристыми лопастями по бокам, перистым «хвостиком» сзади, пятью глазами-пумпочками-на-ножках сверху головы (неизвестно точно – фасеточными или нет) и, в довершение картины, длинным гибким хоботом с расположенной на конце не то зубчатой клешнёй, не то двумя пучками щупалец; никаких других придатков на голове – длинных щупалец, антенн или челюстей – не было. Непарность глаз и хобота как-то плохо вписывается в концепцию билатеральности. Предполагалось даже, что хобот опабинии образовался из слившихся правого и левого ротовых придатков, но нет – он правда исходно был единым, причём подгибался под голову и засовывал еду в расположенный внизу рот, прямо как у слона.
Работа над ошибками
Споры вызвало и строение боковых придатков тела опабинии. Согласно первой реконструкции, выше лежали ребристые лопасти, вероятно, жабры, а под ними – широкие гладкие плавательные. Во второй версии выше располагались широкие ребристые лопасти, а снизу – конические «телескопические» лапки, как у онихофор и тихоходок. В третьей остались только широкие ребристые лопасти, а нижние треугольные стали выступами кишечника.
Потенциальным родственником опабинии может быть Alalcomenaeus cambricus, тоже имевший лопасти по бокам, два больших глаза по сторонам головы и три маленьких по центру, не обладавший антеннами или щупальцами и достоверными ногами.
С большой вероятностью, потомками и ближайшими родственниками керигмачелы и опабинии были Anomalocarididae и очень похожие на них существа. Они тоже были плоскими, с полусегментированным телом, покрытым хитиновыми пластинками, с парными фасеточными глазами на стебельках, со ртом на нижней стороне головы, окружённым диском с пластинчатыми «зубами», рядом с которым крючками загибались членистые ротовые придатки, усаженные шипами. Некоторые аномалокаридиды были вполне приличных размеров – Anamalocaris canadensis и Peytoia nathorsti достигали метра длины, так что они претендуют на роль самых злых хищников своего времени; размеры некоторых ротовых дисков из Чэнцзяна позволили предположить даже двухметровую длину! Известны находки среднекембрийских трилобитов, от которых откушены части, причём размер рта хищника и следы «зубов» максимально подходят к ротовому аппарату аномалокарисов. Обнаружены и очень большие копролиты с остатками трилобитов, а производителями копролитов такого размера, опять же, могли быть только аномалокарисы. С другой стороны, покровы аномалокаридид не были минерализованы, челюсти были не очень твёрды, тогда как трилобиты имели весьма прочный панцирь. Возникает законный вопрос: а могли ли аномалокарисы вообще прокусить кого-то? Чего у аномалокаридид было не отнять, так это размеров и скорости. Возможно, эти преимущества и позволяли им занимать главенствующую роль в раннем и среднем кембрии. Правда, уже в позднем кембрии они стали довольно редки, но протянули вплоть до девона, когда были окончательно побеждены и изведены рыбами.
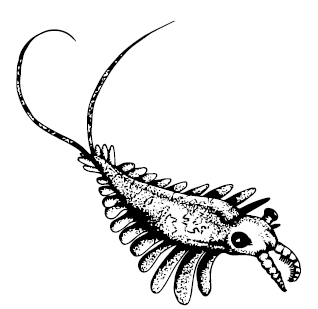
Amplectobelua
Известно довольно много аномалокаридид и родственных им существ. Самые древние – Cassubia infercambriensis – бороздили раннекембрийские моря Польши, наиболее известные и самые большие Anomalocaris и Peytoia найдены по всему миру – в Китае, Польше, Канаде. Два вида Amplectobelua из Китая и Бёрджеса имели особо шипастые ротовые придатки, а Hurdia victoria – мощный головной щит. Pambdelurion whittingtoni из Сириус Пассет обладал «ногами», подобными онихофоровым, а Parapeytoia yunnanensis и Cucumericrus decoratus из Чэнцзяна – даже вполне членистыми почти ногами по краям боковых лопастей, уже очень похожими на ноги членистоногих; ротовые придатки парапейтойи к тому же были вооружены длинными загнутыми на конце подобиями рачьих клешнёй. Те же ротовые придатки у Tamisiocaris borealis были покрыты многочисленными длинными щетинками для фильтрации воды, так что тамисиокарис выступал этаким кембрийским китом.
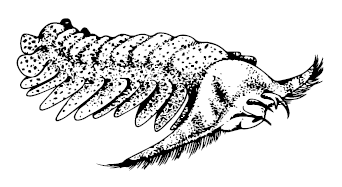
Pambdelurion whittingtoni
Как обычно, мозаика признаков даёт повод для рассуждений: иногда аномалокаридид рассматривают как примитивных членистоногих, но это далеко не факт; согласно альтернативному мнению, они гораздо ближе к тихоходкам.
Работа над ошибками
Сама история открытия аномалокарисов – сплошной палеонтологический детектив. Первыми под названием Anamalocaris canadensis были описаны ротовые придатки, которые были приняты за заднюю часть креветки. Кольцевой рот с пластинками был определен как медузоид «Peytoia nathorsti», а тело – как то ли губка, то ли голотурия «Laggania cambria». В последующем, по мере обнаружения новых отпечатков, головоломка потихоньку собиралась, пока все эти «креветки», «медузы» и «губки» не оказались разными частями одного существа; правда, потом роды Anamalocaris и Peytoia всё же были признаны самостоятельными, но они очень похожи друг на друга.
Подобные проблемы сопровождали и открытие Hurdia victoria: разные части описывались как фрагменты медуз, голотурий, разных членистоногих и аномалокариса, пока не выяснилось, что всё это – элементы одного организма.
* * *Членистоногие Arthropoda появляются в палеонтологической летописи вдруг, внезапно, причём сразу в оптовых количествах. В Чэнцзяне это, например, Naraoia, Leanchoilia и огромное количество других, в Сириус Пассет – трилобитоподобная Campanamuta mantonae и полуметровая Kiisortoqia soperi. Средний кембрий тоже знаменит изобилием нестандартных примитивных членистоногих, их описаны многие десятки: креветкоподобная Yohoia tenuis с двумя коленчатыми четырёхпалыми «руками» спереди и, горизонтальным треугольным плавником сзади, прикрытые двойным панцирем Waptia fieldensis и Plenocaris plena, Odaraia alata с большими глазами на стебельках, двойным панцирем и тремя лопастями на «хвосте», огромные по среднекембрийским меркам двенадцатисантиметровая Sidneyia и девятнадцатисантиметровая расплющенная Helmetia expansa – вероятный предок хелицеровых, совсем уж несуразные рогатые шведские циклопы Cambropachycope clarksoni и Goticaris longispinosa с ногами-вёслами (в Швеции и положено быть всяким троллям). Burgessia bella была похожа на щитня с тонким «хвостом», Molaria spinifera – на мокрицу с длинным непарным членистым «хвостиком», хелицеровый Sanctacaris uncata – на мокрицу с коротким квадратным плоским «хвостиком», Habelia optata была покрыта пупырчатым выпуклым панцирем и тоже заканчивалась «хвостиком», но сделанным из двух шипастых отростков, соединённых вместе. Целый класс Thylacocephala и его родственники – Zhenghecaris, Isoxys, Tuzoia – запаковались в овальные раковинки, из-под которых зыркали пучеглазыми гляделками и высовывали маленькие ножки.
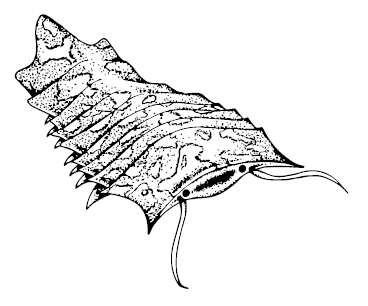
Helmetia expans
Разобраться в этом изобилии нелегко. Существует множество альтернативных схем, предполагающих родство тех или иных групп. На роль примитивнейшего членистоногого претендуют китайские Fuxianhuia protensa, Chengjiangocaris longiformis и Shankouia zhenghei – похожие на мокрицу многоногие твари без каких-то особенных специализаций, но с сердцем, «переднеспинкой», прикрытой крупным щитом, подобием головы и, самое главное, мозгом, совмещающим черты мозга ракообразных, многоножек и насекомых. В среднем кембрии Бёрджеса Tokummia katalepsis представляет следующий уровень усложнения: с двойным панцирем на спине, клешнями спереди, переделанными в пластинчатые жабры задними ногами, а самое главное – верхними и нижними челюстями, сделанными из передних конечностей.
По крайней мере часть кембрийских членистоногих довольно уверенно опознаются как ракообразные, например Canadaspis, несколько тысяч образцов которого найдены в Китае и Канаде; шведские позднекембрийские Agnostus pisiformis и Martinssonia elongata уже совсем креветки креветками. Тогда же появляются такие странные ракообразные, как пятиустки Pentastomida – Heymonsicambria и Haffnericambria из Швеции.
Как вы там, потомки?
Современные ракообразные Crustacea – чрезвычайно разнообразная группа, а потому часто рассматриваются как подтип с несколькими классами. По числу видов ракообразные сильно уступают насекомым, зато намного интереснее морфологически. Особенно удались им вариации конечностей, из которых, оказывается, можно сделать челюсти, ногочелюсти, клешни, ноги, плавники, фильтровальные сеточки и даже жабры.
Современные пятиустки больше похожи на червей, а не раков, паразитируют в дыхательных путях рептилий, птиц и млекопитающих, причём ещё и со сменой хозяев. На ком могли жить кембрийские пятиустки – большой вопрос, так как позвоночных и их дыхательных систем ещё и в помине не было.
Одно из самых колоритных и при том хорошо изученных существ, представленных в Бёрджесе тысячами отпечатков, – Marrella splendens, вместе с несколькими родственниками составляющая особый класс Marrellomorpha. Маррелла больше всего похожа на скелет двухсантиметрового трилобита – тонкое тело с торчащими по бокам тонкими ножками, с длинными усами на голове и двумя парами изогнутых назад «рогов». Покровы марреллы представляли собой дифракционную решётку, благодаря которой зверюшка переливались всеми цветами радуги. Вероятно, маррелла действительно родственна трилобитам.
Трилобиты Trilobita – самые известные существа палеозоя, желанные в коллекции каждого любителя окаменелостей. Их известно от 15 до 50 тысяч видов, так что на планете нет ни одного человека, который бы знал их всех! Изучение трилобитов – отдельная наука, ими можно заниматься всю жизнь и всё равно каждый день удивляться.
В самом начале кембрия трилобиты неизвестны, но, как уже упоминалось, ещё в конце докембрия жила Parvancorina – вероятный предок трилобитов; далее следует прогрессивная линия раннекембрийских мягкотелых родов Primicaris и Skania – Naraoia – Kuamaia – Redlichia, и вот в атдабанском веке Якутии мы встречаем уже древнейшего настоящего трилобита Profallotaspis jakutensis. Как часто бывает, не обошлось и без «живых ископаемых»: в следующем ботомском веке той же Якутии обнаруживается Phytophilaspis pergamena, который вполне мог бы быть предком трилобитов, если бы не жил позже появления этих животных; в Китае подобным пережитком была Cindarella eucala.
Трилобиты уникальны тем, что при едином плане строения (головной щит цефалон с глазами и усиками – членистое тело торакс с утолщением рахисом в центре, выростами плеврами по бокам и ножками снизу – задний щит пигидий) дали фантастическое разнообразие форм. Каждый элемент мог меняться тысячами способов, что сделало трилобитов универсалами – донными и планктонными, хищниками и пылесосами, карликами и гигантами. Первые трилобиты в большинстве были самыми крупными донными хищниками и в раннем и среднем кембрии занимали в этой нише доминирующее положение.
Один из самых стандартных и известных кембрийских трилобитов – Olenoides, в целом похожа на него Damesella paronai, очень большими размерами выделялись Acadoparadoxides briareus – до 45 см – и Paradoxides davidis – до 37 см (а названием – Paradoxides paradoxissimus), Bristolia bristolensis замечательна двумя парами длинных шипов, направленных назад.
Как минимум к концу кембрия некоторые трилобиты развили оригинальные адаптации. Один из самых разнообразных и многочисленных родов Olenus, видимо, вступил в симбиоз с серобактериями. Оленусы жили на дне, где над мутным илом было немного кислорода, а под илом – уже совсем не было. В таких пограничных условиях и трилобиты могли дышать, и серные бактерии заниматься анаэробным хемосинтезом. Тело оленусов неспроста вытянулось: больше сегментов – больше бактерий на дыхательных придатках. Зато в воде с огромным содержанием серы и мизерным содержанием кислорода совсем нет конкурентов и хищников; потому и обнаруживаются оленусы в составе «монокультуры» – фауны из одного вида, в полном видовом одиночестве, зато большими дружными компаниями. Поэтому же они имели очень тонкие панцири – в гиблом болоте оленусов защищаться было не от кого.
Показательно распределение видов трилобитов. Кембрийский океан был холодным в глубине, но тёплым у берега; глубоководные трилобиты без проблем ползали по дну от Китая до Северной Америки, а потому представлены одинаковыми родами и даже видами в крайне удалённых местах, теплолюбивые же мелководные резко отличаются, так как были ограничены в распространении небольшими эндемичными участками.
Одно из главных подразделений членистоногих – подтип хелицеровые Chelicerata. Сейчас к ним относятся классы: паукообразные Arachnida, мечехвосты Merostomata и морские пауки Pycnogonida, из которых успеха достигли лишь первые. В прошлом же группа была гораздо разнообразнее и имела шанс захватить планету.
Маленькая тонкость
Общими легко узнаваемыми чертами всех хелицеровых является отсутствие усиков-антенн, наличие клешнеобразных первых конечностей – хелицер и хватательных вторых – педипальп, разделение тела на головогрудь просому, часто прикрытую крупным головным щитом, и брюшко опистосому, задний конец которой прикрыт ещё одним щитом; тело часто кончается специальным шипообразным сегментом.
В начале кембрия, в атдабанском веке шведский Paleomerus hamiltoni и польский P. makowskii своим овальным членистым телом с несколько увеличенными головным и хвостовым щитами напоминали одновременно и трилобитов, и мечехвостов, и ракоскорпионов; важно, что у них уже были фасеточные глаза. В конце кембрия известен их ближайший родственник Strabops thacheri и более явный родственник мечехвостов – продвинутый Aglaspis spinifer, похожий на трилобита с шипом вместо хвоста.
* * *Конец кембрия, как положено, ознаменовался веянием перемен: появились головоногие моллюски наутилоидеи Nautiloidea, в том числе Plectronoceras, Hunuanoceras, Paleoceras и другие их родичи. Это были ещё совсем маленькие животные, но они были вестниками новой жизни: мчавшиеся, как реактивные пули, в толще воды, они превосходили всех ползающих и колыхавшихся кембрийских тварей, а своим острым клювиком они грозили не только мягкотелым тушкам, но и прочным панцирям. Пахнуло ордовиком…
А где же люди? Первые хордовые
Великое событие кембрия – возникновение вторичноротых и, более того, хордовых животных.
Как известно, билатеральные существа по эмбриологическим данным довольно чётко разделяются на первично- и вторичноротых. Главное их различие – порядок формирования двух концов пищеварительной трубки: у первичноротых на стадии гаструлы сначала впячивается бластопор-рот, а потом открывается анальное отверстие, у вторичноротых – наоборот, сначала анус (причём первоначальный бластопор успевает закрыться, а анус открывается заново примерно там же), а потом – рот. На протяжении двухсот лет зоологи, эмбриологи и анатомы спорили, что бы это значило: является ли человеческая голова аналогом мушиной задницы или головы одинаковы, просто дело в порядке открывания отверстий. Окончательное решение пришло с открытием и изучением Hox-генов (или гомеозисных), которые запускают формирование передне-задней сегментации.
Hox-генов около десятка, причём часто они располагаются на хромосоме по порядку друг за другом. Первые Hox-гены и у людей, и у мух активируют другие группы генов, которые в итоге приводят к образованию головы, следующие – середины тела, последние – заднего конца. Были даже проведены эффектные эксперименты по пересадке Hox-генов от мух мышам и наоборот: несмотря на грандиозное время расхождения эволюционных ветвей, Hox-гены оказались настолько стабильны, что работоспособны при работе в столь отличных чужих организмах. При этом мыши с мушиными генами развивались как надо, с мышиными головами, а мухи с мышиными генами – с мушиными головами. Дело в том, что гены конкретного строения головы – глаз, челюстей, мозга – это уже не Hox-гены, а другие, возникшие позже. Получается, что общий предок не имел деталей строения головы, так что головной конец у первично- и вторичноротых гомологичен (то есть имеет единое эволюционное и эмбриональное происхождение и генетическое обеспечение как раз Hox-генами), а глаза, челюсти и прочее – аналогичны (то есть возникли независимо из разных закладок разными путями в силу схожих потребностей и обеспечиваются разными генами).
Как обычно, существуют довольно многочисленные отклонения и исключения. Так, современная улитка живородка Viviparus неожиданно вторичноротая, хотя все моллюски образцовые первичноротые; щетинкочелюстные Chaetognatha и волосатики Nematomorpha имеют вторичноротый эмбриогенез, но первичноротую анатомию. Для некоторых кольчатых червей, моллюсков, членистоногих и онихофор типична амфистомия, когда исходный единый бластопор вытягивается и разделяется на два отверстия, подобно тому, что предполагается у древнейших предков. У некоторых онихофор и рот, и анус закладываются независимо впереди от бластопора, у немертин тоже независимо, но рот – впереди, а анус – позади от бластопора.
Другое важное отличие – относительное положение в теле пищеварительной и нервной систем: у первичноротых нервные тяжи с ганглиями обычно более-менее парные и идут под кишечником, по брюшной стороне, а у вторичноротых хордовых нервная трубка следует над пищеварительной трубкой по спинной стороне. Перевёрнуты и другие органы: направление тока крови по сосудам, сердце, печень, желток у зародышей, места активации специфических регуляторных белков. Как и с Hox-генами, проводились эксперименты по пересадке генов, определяющих формирование верха и низа у дрозофил («спинной» dpp и «брюшной» sog) и лягушек (chd и bmp-4, соответственно): спинные лягушачьи запускают у мухи образование живота, а брюшные – спины.
Из более специфических отличий нельзя не упомянуть спиральное дробление зиготы и формирование целома из расщепления мезодермы у первичноротых и радиальное дробление и формирование целома из выпячиваний кишечника у вторичноротых.
Есть множество вариантов объяснения явной перевёрнутости вторичноротых.
Самый распространённый гласит, что предки хордовых в некоторый момент перекувырнулись кверху ногами, то есть мы – перевёрнутые червяки. Вообще-то, плавание на спине – довольно обычное дело для животных, питающихся чем-то болтающимся по поверхности воды, достаточно вспомнить клопов-гладышей, щитней, рачков Scapholeberis и улиток Janthina. Если хордовые действительно перевернулись, пара нервных тяжей сблизилась и слилась в трубку. Переворот могли осуществить и донные животные, перешедшие от фильтрования воды растопыренными щупальцами к собиранию пищи с почвы; использовавшийся для ползания членистый хвост мог в итоге превратиться в хвостовой плавник с метамерными мышцами. Между прочим, в почву ланцетники закапываются именно «кверху ногами», то есть нервной трубкой вниз и хордой вверх. Строго говоря, кто вообще определил, где у ланцетника верх, а где низ? Что важнее – положение при плавании или при сидении в грунте?